
Insight into Recent Progress and Perspectives in Improvement of Antioxidant Machinery upon PGPR Augmentation in Plants under Drought Stress: A Review

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Plant Growth Promoting Rhizobacteria Mediated Drought Stress Tolerance in Plants
2.1. Alteration of Host Root System Architecture
2.2. Maintenance of Relative Water Content
2.3. Improvement of Photosynthesis Rate
2.4. Production of Phytohormones
2.4.1. Auxin
2.4.2. Cytokinin
2.4.3. Gibberellin
2.4.4. Abscisic Acid
2.4.5. Salicylic Acid
2.4.6. Jasmonic Acid
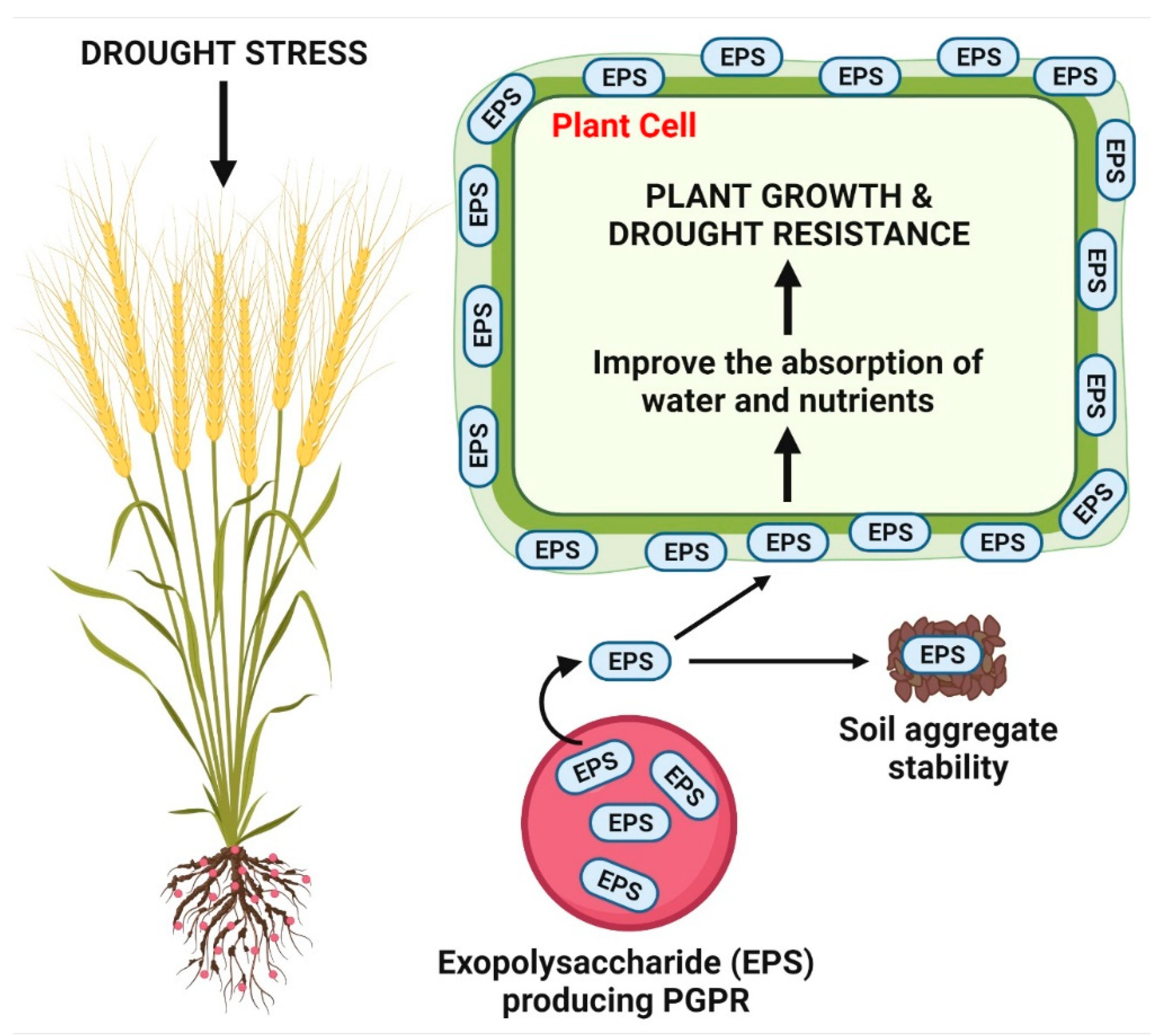
2.5. Production of Exopolysaccharides
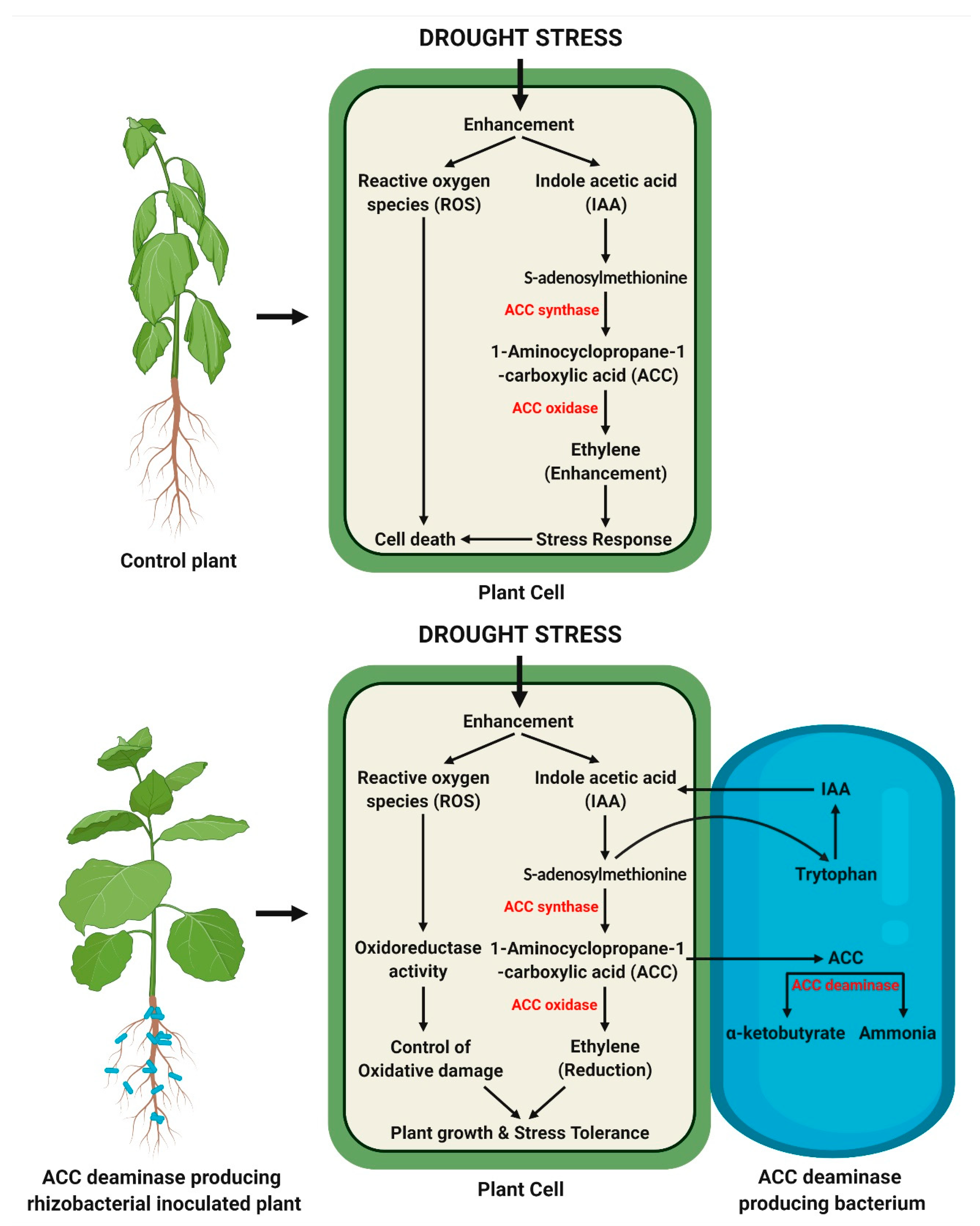
2.6. ACC Deaminase Activity for Reducing Ethylene Levels
2.7. Production of Carotenoids
2.8. Regulation of Emission of Volatiles
2.9. Enhancement of Uptake and Assimilation of Mineral Nutrients
2.10. Siderophore Production
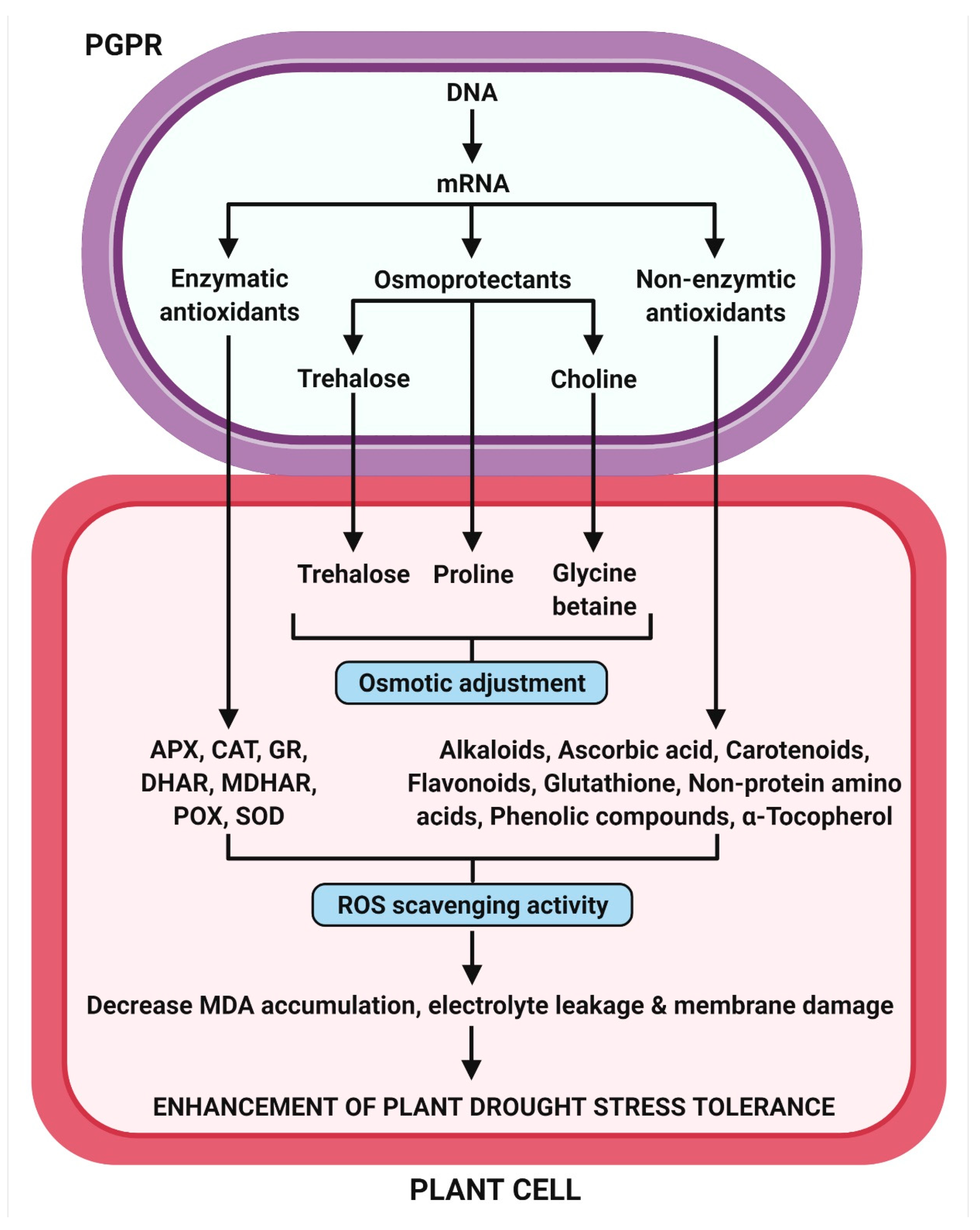
2.11. Induction of Antioxidant Defense Systems
2.12. Reduction in Lipid Peroxidation and Electrolyte Leakage
2.13. Osmotic Adjustment
2.14. Alteration in Expression of Stress Responsive Genes
3. Future Prospects
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ngumbi, E.; Kloepper, J. Bacterial-mediated drought tolerance: Current and future prospects. Appl. Soil Ecol. 2016, 105, 109–125. [Google Scholar] [CrossRef]
- Verma, A.; Shameem, N.; Jatav, H.S.; Sathyanarayana, E.; Parray, J.A.; Poczai, P.; Sayyed, R.Z. Fungal endophytes to combat biotic and abiotic stresses for climate-smart and sustainable agriculture. Front. Plant Sci. 2022, 13, 953836. [Google Scholar] [CrossRef]
- Bhan, S.; Behera, U.K. Conservation agriculture in India–Problems, prospects and policy issues. Int. Soil Water Conserv. Res. 2014, 2, 1–12. [Google Scholar] [CrossRef]
- Ahluwalia, O.; Singh, P.C.; Bhatia, R. A review on drought stress in plants: Implications, mitigation and the role of plant growth promoting rhizobacteria. Resour. Environ. Sustain. 2021, 5, 100032. [Google Scholar] [CrossRef]
- Reshma, P.; Naik, M.K.; Aiyaz, M.; Niranjana, S.R.; Chennappa, G.; Shaikh, S.S.; Sayyed, R.Z. Induced systemic resistance by 2,4-diacetylphloroglucinol positive fluorescent Pseudomonas strains against rice sheath blight. Indian J. Exp. Biol. 2018, 56, 207–212. [Google Scholar]
- Sukmawati, D.; Family, N.; Hidayat, I.; Sayyed, R.Z.; Elsayed, E.A.; Dailin, D.J.; Hanapi, S.Z.; Wadaan, M.A.; Enshasy, H.E. Biocontrol activity of Aureubasidium pullulans and Candida orthopsilosis isolated from Tectona grandis L. phylloplane against Aspergillus sp. in post-harvested citrus fruit. Sustainability 2021, 13, 7479. [Google Scholar] [CrossRef]
- Hamid, B.; Zaman, M.; Farooq, S.; Fatima, S.; Sayyed, R.Z.; Baba, Z.A.; Sheikh, T.A.; Reddy, M.S.; El Enshasy, H.; Gafur, A.; et al. Bacterial plant biostimulants: A sustainable way towards improving growth, productivity, and health of crops. Sustainability 2021, 13, 2856. [Google Scholar] [CrossRef]
- Kour, D.; Kaur, T.; Devi, R.; Yadav, A.; Singh, M.; Joshi, D.; Singh, J.; Suyal, D.C.; Kumar, A.; Rajput, V.D.; et al. Beneficial microbiomes for bioremediation of diverse contaminated environments for environmental sustainability: Present status and future challenges. Environ. Sci. Pollut. Res. Int. 2021, 28, 24917–24939. [Google Scholar] [CrossRef]
- Murali, M.; Gowtham, H.G.; Brijesh Singh, S.; Shilpa, N.; Aiyaz, M.; Niranjana, S.R.; Amruthesh, K.N. Bio-prospecting of ACC deaminase producing rhizobacteria towards sustainable agriculture: A special emphasis on abiotic stress in plants. Appl. Soil Ecol. 2021, 168, 104142. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.S.P. Response of plants to water stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Saleem, A.R.; Brunetti, C.; Khalid, A.; Rocca, G.D.; Raio, A.; Emiliani, G.; Carlo, A.D.; Mahmood, T.; Centritto, M. Drought response of Mucuna pruriens (L.) DC. inoculated with ACC deaminase and IAA producing rhizobacteria. PLoS ONE 2018, 13, e0191218. [Google Scholar] [CrossRef]
- Jabborova, D.; Annapurna, K.; Fayzullaeva, M.; Sulaymonov, K.; Kadirova, D.; Jabbarov, Z.; Sayyed, R.Z. Isolation and characterization of endophytic bacteria from ginger (Zingiber officinale Rosc.). Ann. Phytomed. 2020, 9, 116–121. [Google Scholar] [CrossRef]
- Rajkumar, M.; Ae, N.; Prasad, M.N.V.; Freitas, H. Potential of siderophore-producing bacteria for improving heavy metal phytoextraction. Trends Biotechnol. 2010, 28, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Sayyed, R.Z.; Patel, P.R. Biocontrol potential of siderophore producing heavy metal resistant Alcaligenes sp. and Pseudomonas aeruginosa RZS3 vis-à-vis organophosphorus fungicide. Indian J. Microbiol. 2011, 51, 266–272. [Google Scholar] [CrossRef]
- Ferreira, M.J.; Silva, H.; Cunha, A. Siderophore-producing rhizobacteria as a promising tool for empowering plants to cope with iron limitation in saline soils: A Review. Pedosphere 2019, 29, 409–420. [Google Scholar] [CrossRef]
- Shaikh, S.S.; Wani, S.J.; Sayyed, R.Z. Statistical-based optimization and scale-up of siderophore production process on laboratory bioreactor. 3 Biotech 2016, 6, 69. [Google Scholar] [CrossRef]
- Kalam, S.; Basu, A.; Ahmad, I.; Sayyed, R.Z.; El-Enshasy, H.A.; Dailin, D.J.; Suriani, N.L. Recent understanding of soil Acidobacteria and their ecological significance: A critical review. Front. Microbiol. 2020, 11, 580024. [Google Scholar] [CrossRef] [PubMed]
- Arzanesh, M.H.; Alikhani, H.A.; Khavazi, K.; Rahimian, H.A.; Miransari, M. Wheat (Triticum aestivum L.) growth enhancement by Azospirillum sp. under drought stress. World J. Microbiol. Biotechnol. 2011, 27, 197–205. [Google Scholar] [CrossRef]
- Patel, P.R.; Shaikh, S.S.; Sayyed, R.Z. Modified chrome azurol S method for detection and estimation of siderophores having affinity for metal ions other than iron. Environ. Sustain. 2018, 1, 81–87. [Google Scholar] [CrossRef]
- Sayyed, R.Z.; Seifi, S.; Patel, P.R.; Shaikh, S.S.; Jadhav, H.P.; El Enshasy, H. Siderophore production in groundnut rhizosphere isolate, Achromobacter sp. RZS2 influenced by physicochemical factors and metal ions. Environ. Sustain. 2019, 2, 117–124. [Google Scholar] [CrossRef]
- Cohen, A.C.; Bottini, R.; Pontin, M.; Berli, F.J.; Moreno, D.; Boccanlandro, H.; Travaglia, C.N.; Piccoli, P.N. Azospirillum brasilense ameliorates the response of Arabidopsis thaliana to drought mainly via enhancement of ABA levels. Physiol. Plant. 2015, 153, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Jabborova, D.; Wirth, S.; Kannepalli, A.; Narimanov, A.; Desouky, S.; Davranov, K.; Sayyed, R.Z.; El Enshasy, H.; Malek, R.A.; Syed, A.; et al. Co-inoculation of rhizobacteria and biochar application improves growth and nutrients in Soybean and enriches soil nutrients and enzymes. Agronomy 2020, 10, 1142. [Google Scholar] [CrossRef]
- Marulanda, A.; Barea, J.M.; Azcón, R. Stimulation of plant growth and drought tolerance by native microorganisms (AM fungi and bacteria) from dry environments: Mechanisms related to bacterial effectiveness. J. Plant Growth Regul. 2009, 28, 115–124. [Google Scholar] [CrossRef]
- Huang, X.F.; Zhou, D.; Lapsansky, E.R.; Reardon, K.F.; Guo, J.; Andales, M.J.; Vivanco, J.M.; Manter, D.K. Mitsuaria sp. and Burkholderia sp. from Arabidopsis rhizosphere enhance drought tolerance in Arabidopsis thaliana and maize (Zea mays L.). Plant Soil 2017, 419, 523–539. [Google Scholar] [CrossRef]
- Jochum, M.D.; McWilliams, K.L.; Borrego, E.J.; Kolomiets, M.V.; Niu, G.; Pierson, E.A.; Jo, Y.K. Bioprospecting plant growth-promoting rhizobacteria that mitigate drought stress in grasses. Front. Microbiol. 2019, 10, 2106. [Google Scholar] [CrossRef]
- Silva, E.R.; Zoz, J.; Oliveira, C.E.S.; Zuffo, A.M.; Steiner, F.; Zoz, T.; Vendruscolo, E.P. Can co-inoculation of Bradyrhizobium and azospirillum alleviate adverse effects of drought stress on soybean (Glycine max L. Merrill.)? Arch. Microbiol. 2019, 201, 325–335. [Google Scholar] [CrossRef]
- Nithyapriya, S.; Lalitha, S.; Sayyed, R.Z.; Reddy, M.S.; Dailin, D.J.; El Enshasy, H.A.; LuhSuriani, N.; Herlambang, S. Production, purification, and characterization of Bacillibactin siderophore of Bacillus subtilis and its application for improvement in plant growth and oil content in Sesame. Sustainability 2021, 13, 5394. [Google Scholar] [CrossRef]
- Farooq, M.; Hussain, M.; Wahid, A.; Siddique, K.H.M. Drought stress in plants: An over view. In Plant Responses to Drought Stress; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 1–33. [Google Scholar] [CrossRef]
- Castillo, P.; Escalante, M.; Gallardo, M.; Alemano, S.; Abdala, G. Effects of bacterial single inoculation and co-inoculation on growth and phytohormone production of sunflower seedlings under water stress. Acta Physiol. Plant. 2013, 35, 2299–2309. [Google Scholar] [CrossRef]
- García, J.E.; Maroniche, G.; Creus, C.; Suárez-Rodríguez, R.; Ramirez-Trujillo, J.A.; Groppa, M.D. In Vitro PGPR properties and osmotic tolerance of different Azospirillum native strains and their effects on growth of maize under drought stress. Microbiol. Res. 2017, 202, 21–29. [Google Scholar] [CrossRef]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; El Enshasy, H. Plant Growth Promoting Rhizobacteria (PGPR) as green bioinoculants: Recent developments, constraints, and prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Khan, N.; Bano, A. Exopolysaccharide producing rhizobacteria and their impact on growth and drought tolerance of wheat grown under rainfed conditions. PLoS ONE 2019, 14, e0222302. [Google Scholar] [CrossRef]
- Kudoyarova, G.; Arkhipova, T.; Korshunova, T.; Bakaeva, M.; Loginov, O.; Dodd, I.C. Phytohormone mediation of interactions between plants and non-symbiotic growth promoting bacteria under edaphic stresses. Front. Plant Sci. 2019, 10, 1368. [Google Scholar] [CrossRef]
- Gowtham, H.G.; Duraivadivel, P.; Ayusman, S.; Sayani, D.; Gholap, S.L.; Niranjana, S.R.; Hariprasad, P. ABA analogue produced by Bacillus marisflavi modulates the physiological response of Brassica juncea L. under drought stress. Appl. Soil Ecol. 2021, 159, 103845. [Google Scholar] [CrossRef]
- Duan, B.; Li, L.; Chen, G.; Su-Zhou, C.; Li, Y.; Merkeryan, H.; Liu, W.; Liu, X. 1-Aminocyclopropane-1-carboxylate deaminase-producing plant growth-promoting rhizobacteria improve drought stress tolerance in Grapevine (Vitis vinifera L.). Front. Plant Sci. 2021, 12, 706990. [Google Scholar] [CrossRef]
- Nadeem, S.M.; Ahmad, M.; Tufail, M.A.; Asghar, H.N.; Nazli, F.; Zahir, Z.A. Appraising the potential of EPS-producing rhizobacteria with ACC-deaminase activity to improve growth and physiology of maize under drought stress. Physiol. Plant. 2020, 172, 463–476. [Google Scholar] [CrossRef]
- Timmusk, S.; Abd El-Daim, I.A.; Copolovici, L.; Tanilas, T.; Kännaste, A.; Behers, L.; Nevo, E.; Seisenbaeva, G.; Stenström, E.; Niinemets, Ü. Drought-tolerance of wheat improved by rhizosphere bacteria from harsh environments: Enhanced biomass production and reduced emissions of stress volatiles. PLoS ONE 2014, 9, e96086. [Google Scholar] [CrossRef]
- Martins, S.J.; Rocha, G.A.; de Melo, H.C.; Georg, R.D.C.; Ulhôa, C.J.; Dianese, É.D.C.; Oshiquiri, L.H.; da Cunha, M.G.; da Rocha, M.R.; de Araújo, L.G.; et al. Plant-associated bacteria mitigate drought stress in soybean. Environ. Sci. Pollut. Res. 2018, 25, 13676–13686. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Ma, H.; Peng, L.; Du, Z.; Ma, B.; Liu, X. Effect of the inoculation of plant growth-promoting rhizobacteria on the photosynthetic characteristics of Sambucus williamsii Hance container seedlings under drought stress. AMB Expr. 2019, 9, 169. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd Allah, E.F.; Hashem, A. Phytohormones and beneficial microbes: Essential components for plants to balance stress and fitness. Front. Microbiol. 2017, 8, 2104. [Google Scholar] [CrossRef] [PubMed]
- Hariprasad, P.; Gowtham, H.G.; Gourav, C. Beneficial plant-associated bacteria modulate host hormonal system enhancing plant resistance toward abiotic stress. In Biocontrol Agents and Secondary Metabolites; Jogaiah, S., Ed.; Woodhead Publishing: Kidlington, UK, 2021; pp. 113–151. [Google Scholar] [CrossRef]
- Khan, I.; Awan, S.A.; Ikram, R.; Rizwan, M.; Akhtar, N.; Yasmin, H.; Sayyed, R.Z.; Ali, S.; Ilyas, N. Effects of 24-epibrassinolide on plant growth, antioxidants defense system, and endogenous hormones in two wheat varieties under drought stress. Physiol. Plant. 2021, 172, 696–706. [Google Scholar] [CrossRef]
- Hayat, R.; Ali, S.; Amara, U.; Khalid, R.; Ahmed, I. Soil beneficial bacteria and their role in plant growth promotion: A review. Ann. Microbiol. 2010, 60, 579–598. [Google Scholar] [CrossRef]
- Khan, N.; Ali, S.; Shahid, M.A.; Mustafa, A.; Sayyed, R.Z.; Curá, J.A. Insights into the interactions among roots, rhizosphere, and rhizobacteria for improving plant growth and tolerance to abiotic stresses: A review. Cells 2021, 10, 1551. [Google Scholar] [CrossRef]
- Raheem, A.; Shaposhnikov, A.; Belimov, A.A.; Dodd, I.C.; Ali, B. Auxin production by rhizobacteria was associated with improved yield of wheat (Triticum aestivum L.) under drought stress. Arch. Agron. Soil Sci. 2018, 64, 574–587. [Google Scholar] [CrossRef]
- Uzma, M.; Iqbal, A.; Hasnain, S. Drought tolerance induction and growth promotion by indole acetic acid producing Pseudomonas aeruginosa in Vigna radiate. PLoS ONE 2022, 17, e0262932. [Google Scholar] [CrossRef]
- Arkhipova, T.N.; Prinsen, E.; Veselov, S.U.; Martinenko, E.V.; Melentiev, A.I.; Kudoyarova, G.R. Cytokinin producing bacteria enhance plant growth in drying soil. Plant Soil 2007, 292, 305–315. [Google Scholar] [CrossRef]
- Liu, F.; Xing, S.; Ma, H.; Du, Z.; Ma, B. Cytokinin-producing, plant growth-promoting rhizobacteria that confer resistance to drought stress in Platycladusorientalis container seedlings. Appl. Microbiol. Biotechnol. 2013, 97, 9155–9164. [Google Scholar] [CrossRef] [PubMed]
- Gowtham, H.G.; Murali, M.; Brijesh Singh, S.; Lakshmeesha, T.R.; Murthy, K.; Amruthesh, K.N.; Niranjana, S.R. Plant growth promoting rhizobacteria- Bacillus amyloliquefaciens improves plant growth and induces resistance in chilli against anthracnose disease. Biol. Control 2018, 126, 209–217. [Google Scholar] [CrossRef]
- Kang, S.M.; Radhakrishnan, R.; Khan, A.L.; Kim, M.J.; Park, J.M.; Kim, B.R.; Shin, D.H.; Lee, I.J. Gibberellin secreting rhizobacterium, Pseudomonas putida H-2-3 modulates the hormonal and stress physiology of soybean to improve the plant growth under saline and drought conditions. Plant Physiol. Biochem. 2014, 84, 115–124. [Google Scholar] [CrossRef]
- Salomon, M.V.; Bottini, R.; de Souza Filho, G.A.; Cohen, A.C.; Moreno, D.; Gil, M.; Piccoli, P. Bacteria isolated from roots and rhizosphere of Vitis vinifera retard water losses, induce abscisic acid accumulation and synthesis of defense-related terpenes in in vitro cultured grapevine. Physiol. Plant. 2014, 151, 359–374. [Google Scholar] [CrossRef]
- Gowtham, H.G.; Duraivadivel, P.; Hariprasad, P.; Niranjana, S.R. A novel split-pot bioassay to screen indole acetic acid producing rhizobacteria for the improvement of plant growth in tomato [Solanum lycopersicum L.]. Sci. Hortic. 2017, 224, 351–357. [Google Scholar] [CrossRef]
- Farber, M.; Attia, Z.; Weiss, D. Cytokinin activity increases stomatal density and transpiration rate in tomato. J. Exp. Bot. 2016, 67, 6351–6362. [Google Scholar] [CrossRef]
- Al Mahmud, J.; Biswas, P.K.; Nahar, K.; Fujita, M.; Hasanuzzaman, M. Exogenous application of gibberellic acidmitigates drought-induced damage inspring wheat. Acta Agrobot. 2019, 72, 1776. [Google Scholar] [CrossRef]
- Joo, G.J.; Kim, Y.M.; Kim, J.T.; Rhee, I.K.; Kim, J.H.; Lee, I.J. Gibberellins-producing rhizobacteria increase endogenous gibberellins content and promote growth of red peppers. J. Microbiol. 2005, 43, 510–515. [Google Scholar]
- Kang, S.M.; Khan, A.L.; Hamayun, M.; Hussain, J.; Joo, G.J.; You, Y.H.; Kim, J.G.; Lee, I.J. Gibberellin-producing Promicromonospora sp. SE188 improves Solanum lycopersicum plant growth and influences endogenous plant hormones. J. Microbiol. 2012, 50, 902–909. [Google Scholar] [CrossRef]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought-from genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef]
- Muhammad Aslam, M.; Waseem, M.; Jakada, B.H.; Okal, E.J.; Lei, Z.; Saqib, H.S.A.; Yuan, W.; Xu, W.; Zhang, Q. Mechanisms of Abscisic acid-mediated drought stress responses in plants. Int. J. Mol. Sci. 2022, 23, 1084. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.R.; Fatma, M.; Per, T.S.; Anjum, N.A.; Khan, N.A. Salicylic acid-induced abiotic stress tolerance and underlying mechanisms in plants. Front. Plant Sci. 2015, 6, 462. [Google Scholar] [CrossRef]
- Ali, S.A.M.; Sayyed, R.Z.; Mir, M.I.; Khan, M.Y.; Hameeda, B.; Alkhanani, M.F.; Haque, S.; Mohammad Al Tawaha, A.R.; Poczai, P. Induction of systemic resistance in maize and antibiofilm activity of surfactin from Bacillus velezensis MS20. Front. Microbiol. 2022, 13, 879739. [Google Scholar] [CrossRef]
- Bakker, P.A.H.M.; Ran, L.; Mercado-Blanco, J. Rhizobacterial salicylate production provokes headaches! Plant Soil 2014, 382, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.S.; Baek, K.H. Jasmonic acid signaling pathway in response to abiotic stresses in plants. Int. J. Mol. Sci. 2020, 21, 621. [Google Scholar] [CrossRef]
- Forchetti, G.; Masciarelli, O.; Alemano, S.; Alvarez, D.; Abdala, G. Endophytic bacteria in sunflower (Helianthus annuus L.): Isolation, characterization, and production of jasmonates and abscisic acid in culture medium. Appl. Microbiol. Biotechnol. 2007, 76, 1145–1152. [Google Scholar] [CrossRef]
- Sayyed, R.Z.; Patel, P.R.; Shaikh, S.S. Plant growth promotion and root colonization by EPS producing Enterobacter sp. RZS5 under heavy metal contaminated soil. Indian J. Exp. Biol. 2015, 53, 116–123. [Google Scholar] [PubMed]
- Ilyas, N.; Mumtaz, K.; Akhtar, N.; Yasmin, H.; Sayyed, R.Z.; Khan, W.; Enshasy, H.A.E.; Dailin, D.J.; Elsayed, E.A.; Ali, Z. Exopolysaccharides producing bacteria for the amelioration of drought stress in Wheat. Sustainability 2020, 12, 8876. [Google Scholar] [CrossRef]
- Naseem, H.; Ahsan, M.; Shahid, M.A.; Khan, N. Exopolysaccharides producing rhizobacteria and their role in plant growth and drought tolerance. J. Basic Microbiol. 2018, 58, 1009–1022. [Google Scholar] [CrossRef]
- Ansari, F.A.; Jabeen, M.; Ahmad, I. Pseudomonas azotoformans FAP5, a novel biofilm-forming PGPR strain, alleviates drought stress in wheat plant. Int. J. Environ. Sci. Technol. 2021, 18, 3855–3870. [Google Scholar] [CrossRef]
- Alami, Y.; Achouak, W.; Marol, C.; Heulin, T. Rhizosphere soil aggregation and plant growth promotion of sunflowers by an exopolysaccharide-producing Rhizobium sp. strain isolated from sunflower roots. Appl. Environ. Microb. 2000, 66, 3393–3398. [Google Scholar] [CrossRef]
- Niu, X.; Song, L.; Xiao, Y.; Ge, W. Drought-tolerant plant growth-promoting rhizobacteria associated with Foxtail millet in a semi-arid agroecosystem and their potential in alleviating drought stress. Front. Microbiol. 2018, 8, 2580. [Google Scholar] [CrossRef]
- Vardharajula, S.; Ali, S.Z.; Grover, M.; Reddy, G.; Bandi, V. Drought-tolerant plant growth promoting Bacillus spp.: Effect on growth, osmolytes, and antioxidant status of maize under drought stress. J. Plant Interact. 2011, 6, 1–14. [Google Scholar] [CrossRef]
- Naseem, H.; Bano, A. Role of plant growth-promoting rhizobacteria and their exopolysaccharide in drought tolerance of maize. J. Plant Interact. 2014, 9, 689–701. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, D.; Gupta, A.; Mohapatra, S. A comparative analysis of exopolysaccharide and phytohormone secretions by four drought-tolerant rhizobacterial strains and their impact on osmotic-stress mitigation in Arabidopsis thaliana. World J. Microbiol. Biotechnol. 2019, 35, 90. [Google Scholar] [CrossRef]
- Iqbal, N.; Khan, N.A.; Ferrante, A.; Trivellini, A.; Francini, A.; Khan, M.I.R. Ethylene role in plant growth, development and senescence: Interaction with other phytohormones. Front. Plant Sci. 2017, 8, 475. [Google Scholar] [CrossRef]
- Glick, B.R. Bacteria with ACC deaminase can promote plant growth and help to feed the world. Microbiol. Res. 2014, 169, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Sagar, A.; Sayyed, R.Z.; Ramteke, P.W.; Sharma, S.; Marraiki, N.; Elgorban, A.M.; Syed, A. ACC deaminase and antioxidant enzymes producing halophilic Enterobacter sp. PR14 promotes the growth of rice and millets under salinity stress. Physiol. Mol. Biol. Plants 2020, 26, 1847–1854. [Google Scholar] [CrossRef]
- Brijesh Singh, S.; Gowtham, H.G.; Murali, M.; Hariprasad, P.; Lakshmeesha, T.R.; Narasimha Murthy, K.; Amruthesh, K.N.; Niranjana, S.R. Plant growth promoting ability of ACC deaminase producing rhizobacteria native to Sunflower (Helianthus annuus L.). Biocatal. Agric. Biotechnol. 2019, 18, 101089. [Google Scholar] [CrossRef]
- Gowtham, H.G.; Brijesh Singh, S.; Murali, M.; Shilpa, N.; Prasad, M.; Aiyaz, M.; Amruthesh, K.N.; Niranjana, S.R. Induction of drought tolerance in tomato upon the application of ACC deaminase producing plant growth promoting rhizobacterium Bacillus subtilis Rhizo SF 48. Microbiol. Res. 2020, 234, 126422. [Google Scholar] [CrossRef]
- Saikia, J.; Sarma, R.K.; Dhandia, R.; Yadav, A.; Bharali, R.; Gupta, V.K.; Saikia, R. Alleviation of drought stress in pulse crops with ACC deaminase producing rhizobacteria isolated from acidic soil of Northeast India. Sci. Rep. 2018, 8, 3560. [Google Scholar] [CrossRef]
- Chandra, D.; Srivastava, R.; Gupta, V.V.S.R.; Franco, C.M.M.; Sharma, A.K. Evaluation of ACC-deaminase-producing rhizobacteria to alleviate water-stress impacts in wheat (Triticum aestivum L.) plants. Can. J. Microbiol. 2019, 65, 387–403. [Google Scholar] [CrossRef]
- Murali, M.; Brijesh Singh, S.; Gowtham, H.G.; Shilpa, N.; Prasad, M.; Aiyaz, M.; Amruthesh, K.N. Induction of drought tolerance in Pennisetum glaucum by ACC deaminase producing PGPR- Bacillus amyloliquefaciens through antioxidant defense system. Microbiol. Res. 2021, 253, 126891. [Google Scholar] [CrossRef]
- Barnawal, D.; Maji, D.; Bharti, N.; Chanotiya, C.S.; Kalra, A. ACC deaminase-containing Bacillus subtilis reduces stress ethylene-induced damage and improves mycorrhizal colonization and rhizobial nodulation in Trigonella foenum-graecum under drought stress. J. Plant Growth Regul. 2013, 32, 809–822. [Google Scholar] [CrossRef]
- Belimov, A.A.; Dodd, I.C.; Safronova, V.I.; Shaposhnikov, A.I.; Azarova, T.S.; Makarova, N.M.; Davies, W.J.; Tikhonovich, I.A. Rhizobacteria that produce auxins and contain 1-amino-cyclopropane-1-carboxylic acid deaminase decrease amino acid concentrations in the rhizosphere and improve growth and yield of well-watered and water-limited potato (Solanum tuberosum). Ann. Appl. Biol. 2015, 167, 11–25. [Google Scholar] [CrossRef]
- Vurukonda, S.S.K.P.; Vardharajula, S.; Shrivastava, M.; SkZ, A. Multifunctional Pseudomonas putida strain FBKV2 from arid rhizosphere soil and its growth promotional effects on maize under drought stress. Rhizosphere 2016, 1, 4–13. [Google Scholar] [CrossRef]
- Chandra, D.; Srivastava, R.; Sharma, A.K. Influence of IAA and ACC deaminase producing fluorescent Pseudomonads in alleviating drought stress in Wheat (Triticum aestivum). Agric. Res. 2018, 7, 290–299. [Google Scholar] [CrossRef]
- Singh, S.B.; Gowtham, H.G.; Aiyaz, M.; Niranjana, S.R. Changes in enzymatic and non-enzymatic defence systems induced by ACCd producing PGPR aid sunflower plants to tolerate drought stress. Int. J. Pharm. Biol. Sci. 2019, 9, 782–791. [Google Scholar]
- Chandra, D.; Srivastava, R.; Glick, B.R.; Sharma, A.K. Rhizobacteria producing ACC deaminase mitigate water-stress response in finger millet (Eleusine coracana (L.) Gaertn.). 3 Biotech 2020, 10, 65. [Google Scholar] [CrossRef]
- Zarei, T.; Moradi, A.; Kazemeini, S.A.; Akhgar, A.; Rahi, A.A. The role of ACC deaminase producing bacteria in improving sweet corn (Zea mays L. var saccharata) productivity under limited availability of irrigation water. Sci. Rep. 2020, 10, 20361. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, L.; Hao, R.; Bai, X.; Wang, Y.; Yu, X. Drought-tolerant plant growth-promoting rhizobacteria isolated from jujube (Ziziphus jujuba) and their potential to enhance drought tolerance. Plant Soil 2020, 452, 423–440. [Google Scholar] [CrossRef]
- Brunetti, C.; Saleem, A.R.; Rocca, G.D.; Emiliani, G.; Carlo, A.D.; Balestrini, R.; Khalid, A.; Mahmood, T.; Centritto, M. Effects of plant growth-promoting rhizobacteria strains producing ACC deaminase on photosynthesis, isoprene emission, ethylene formation and growth of Mucuna pruriens (L.) DC. in response to water deficit. J. Biotechnol. 2021, 331, 53–62. [Google Scholar] [CrossRef]
- Sandhya, V.; SK, Z.A.; Grover, M.; Reddy, G.; Venkateswarlu, B. Alleviation of drought stress effects in sunflower seedlings by the exopolysaccharides producing Pseudomonas putida strain GAP-P45. Biol. Fertil. Soils 2009, 46, 17–26. [Google Scholar] [CrossRef]
- Cho, S.M.; Kang, B.R.; Han, S.H.; Anderson, A.J.; Park, J.Y.; Lee, Y.H.; Cho, B.H.; Yang, K.Y.; Ryu, C.M.; Kim, Y.C. 2R,3R-Butanediol, a bacterial volatile produced by Pseudomonas chlororaphis O6, is involved in induction of systemic tolerance to drought in Arabidopsis thaliana. Mol. Plant Microb. Interact. 2008, 21, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Uarrota, V.G.; Stefen, D.L.V.; Leolato, L.S.; Gindri, D.M.; Nerling, D. Revisiting carotenoids and their role in plant stress responses: From biosynthesis to plant signaling mechanisms during stress. In Antioxidants and Antioxidant Enzymes in Higher Plants; Gupta, D., Palma, J., Corpas, F., Eds.; Springer: Cham, Switzerland, 2018; pp. 207–232. [Google Scholar] [CrossRef]
- Fallah, M.; Hadi, H.; Amirnia, R.; Hassanzadeh-Ghorttapeh, A.; Zuan, A.T.K.; Sayyed, R.Z. Eco-friendly soil amendments improve growth, antioxidant activities, and root colonization in lingrain (Linum usitatissimum L.) under drought conditions. PLoS ONE 2021, 16, e0261225. [Google Scholar] [CrossRef]
- Polapally, R.; Mansani, M.; Rajkumar, K.; Burgula, S.; Hameeda, B.; Alhazmi, A.; Bantun, F.; Almalki, A.H.; Haque, S.; El Enshasy, H.A.; et al. Melanin pigment of Streptomyces puniceus RHPR9 exhibits antibacterial, antioxidant and anticancer activities. PLoS ONE 2022, 17, e0266676. [Google Scholar] [CrossRef]
- Nisar, N.; Li, L.; Lu, S.; Khin, N.C.; Pogson, B.J. Carotenoid metabolism in plants. Mol. Plant 2015, 8, 68–82. [Google Scholar] [CrossRef] [PubMed]
- Yasmin, H.; Nosheen, A.; Naz, R.; Bano, A.; Keyani, R. L-tryptophan-assisted PGPR-mediated induction of drought tolerance in maize (Zea mays L.). J. Plant Interact. 2017, 12, 567–578. [Google Scholar] [CrossRef]
- Batool, T.; Ali, S.; Seleiman, M.F.; Naveed, N.H.; Ali, A.; Ahmed, K.; Abid, M.; Rizwan, M.; Shahid, M.R.; Alotaibi, M.; et al. Plant growth promoting rhizobacteria alleviates drought stress in potato in response to suppressive oxidative stress and antioxidant enzymes activities. Sci. Rep. 2020, 10, 16975. [Google Scholar] [CrossRef]
- Erb, M. Volatiles as inducers and suppressors of plant defense and immunity—Origins, specificity, perception and signaling. Curr. Opin. Plant Biol. 2018, 44, 117–121. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Bravo, L.A.; Copolovici, L. Changes in photosynthetic rate and stress volatile emissions through desiccation-rehydration cycles in desiccation-tolerant epiphytic filmy ferns (Hymenophyllaceae). Plant Cell Environ. 2018, 41, 1605–1617. [Google Scholar] [CrossRef]
- Bomle, D.V.; Kiran, A.; Kumar, J.K.; Nagaraj, L.S.; Pradeep, C.K.; Ansari, M.A.; Alghamdi, S.; Kabrah, A.; Assaggaf, H.; Dablool, A.S.; et al. Plants saline environment in perception with rhizosphere bacteria containing 1-aminocyclopropane-1-carboxylate deaminase. Int. J. Mol. Sci. 2021, 22, 11461. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.M.; Zhang, H. The effects of bacterial volatile emissions on plant abiotic stress tolerance. Front. Plant Sci. 2015, 6, 774. [Google Scholar] [CrossRef]
- Rouphael, Y.; Cardarelli, M.; Schwarz, D.; Franken, P.; Colla, G. Effects of drought on nutrient uptake and assimilation in vegetable crops. In Plant Responses to Drought Stress; Aroca, R., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 171–195. [Google Scholar] [CrossRef]
- Baba, Z.A.; Hamid, B.; Sheikh, T.A.; Alotaibi, S.H.; El Enshasy, H.A.; Ansari, M.J.; Zuan, A.T.K.; Sayyed, R.Z. Psychrotolerant Mesorhizobium sp. isolated from temperate and cold desert regions solubilizes potassium and produces multiple plant growth promoting metabolites. Molecules 2021, 26, 5758. [Google Scholar] [CrossRef] [PubMed]
- Ahanger, M.A.; Morad-Talab, N.; Abd-Allah, E.F.; Ahmad, P.; Hajiboland, R. Plant growth under drought stress. In Water Stress and Crop Plants: A Sustainable Approach; Ahmad, P., Ed.; John Wiley & Sons: West Sussex, UK, 2016; pp. 649–668. [Google Scholar] [CrossRef]
- Kapadia, C.; Sayyed, R.Z.; El Enshasy, H.A.; Vaidya, H.; Sharma, D.; Patel, N.; Malek, R.A.; Syed, A.; Elgorban, A.M.; Ahmad, K.; et al. Halotolerant microbial consortia for sustainable mitigation of salinity stress, growth promotion, and mineral uptake in tomato plants and soil nutrient enrichment. Sustainability 2021, 13, 8369. [Google Scholar] [CrossRef]
- Armada, E.; Leite, M.F.A.; Medina, A.; Azcón, R.; Kuramae, E.E. Native bacteria promote plant growth under drought stress condition without impacting the rhizomicrobiome. FEMS Microbiol. Ecol. 2018, 94, fiy092. [Google Scholar] [CrossRef]
- Jabborova, D.; Sayyed, R.Z.; Azimov, A.; Jabbarov, Z.; Matchanov, A.; Enakiev, Y.; Baazeem, A.; El Sabagh, A.; Danish, S.; Datta, R. Impact of mineral fertilizers on mineral nutrients in the ginger rhizome and on soil enzymes activities and soil properties. Saudi J. Biol Sci. 2021, 28, 5268–5274. [Google Scholar] [CrossRef]
- Jabborova, D.; Sulaymanov, K.; Sayyed, R.Z.; Alotaibi, S.H.; Enakiev, Y.; Azimov, A.; Jabbarov, Z.; Ansari, M.J.; Fahad, S.; Danish, S.; et al. Mineral fertilizers improves the quality of turmeric and soil. Sustainability 2021, 13, 9437. [Google Scholar] [CrossRef]
- Khumairah, F.H.; Setiawati, M.R.; Fitriatin, B.N.; Simarmata, T.; Alfaraj, S.; Ansari, M.J.; Enshasy, H.A.E.; Sayyed, R.Z.; Najafi, S. Halotolerant Plant growth-promoting rhizobacteria isolated from saline soil improve nitrogen fixation and alleviate salt stress in rice plants. Front. Microbiol. 2022, 13, 905210. [Google Scholar] [CrossRef]
- Cruz de Carvalho, M.H. Drought stress and reactive oxygen species: Production, scavenging and signaling. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Kusale, S.P.; Attar, Y.C.; Sayyed, R.Z.; Malek, R.A.; Ilyas, N.; Suriani, N.L.; Khan, N.; El Enshasy, H.A. Production of plant beneficial and antioxidants metabolites by Klebsiella variicola under salinity stress. Molecules 2021, 26, 1894. [Google Scholar] [CrossRef] [PubMed]
- Najafi, S.; Nazari Nasi, H.; Tuncturk, R.; Tuncturk, M.; Sayyed, R.Z.; Amirnia, R. Biofertilizer application enhances drought stress tolerance and alters the antioxidant enzymes in medicinal pumpkin (Cucurbita pepo convar. pepo var. Styriaca). Horticulturae 2021, 7, 588. [Google Scholar] [CrossRef]
- Sandhya, V.; Ali, S.Z.; Grover, M.; Reddy, G.; Venkateswarlu, B. Effect of plant growth promoting Pseudomonas spp. on compatible solutes, antioxidant status and plant growth of maize under drought stress. Plant Growth Regul. 2010, 62, 21–30. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Ghorbanpour, M.; Hatami, M.; Khavazi, K. Role of plant growth promoting rhizobacteria on antioxidant enzyme activities andtropane alkaloid production of Hyoscyamus niger under water deficit stress. Turk. J. Biol. 2013, 37, 350–360. [Google Scholar]
- Chiappero, J.; Cappellari, L.D.R.; Alderete, L.G.S.; Palermo, T.B.; Banchio, E. Plant growth promoting rhizobacteria improve the antioxidant status in Mentha piperita grown under drought stress leading to an enhancement of plant growth and total phenolic content. Ind. Crops Prod. 2019, 139, 111553. [Google Scholar] [CrossRef]
- Abdelaal, K.; AlKahtani, M.; Attia, K.; Hafez, Y.; Király, L.; Künstler, A. The role of plant growth-promoting bacteria in alleviating the adverse effects of drought on plants. Biology 2021, 10, 520. [Google Scholar] [CrossRef]
- Asghari, B.; Khademian, R.; Sedaghati, B. Plant growth promoting rhizobacteria (PGPR) confer drought resistance and stimulate biosynthesis of secondary metabolites in pennyroyal (Mentha pulegium L.) under water shortage condition. Sci. Hortic. 2020, 263, 109132. [Google Scholar] [CrossRef]
- Turner, N.C. Turgor maintenance by osmotic adjustment, an adaptive mechanism for coping with plant water deficits. Plant Cell Environ. 2017, 40, 1–3. [Google Scholar] [CrossRef]
- Arora, H.; Sharma, A.; Poczai, P.; Sharma, S.; Haron, F.F.; Gafur, A.; Sayyed, R.Z. Plant-derived protectants in combating soil-borne fungal infections in tomato and chilli. J. Fungi 2022, 8, 213. [Google Scholar] [CrossRef]
- Sheikh, T.; Hamid, B.; Baba, Z.; Iqbal, S.; Yatoo, A.; Fatima, S.; Nabi, A.; Kanth, R.; Dar, K.; Hussain, N.; et al. Extracellular polymeric substances in psychrophilic cyanobacteria: A potential bioflocculant and carbon sink to mitigate cold stress. Biocatal. Agric. Biotechnol. 2022, 42, 102375. [Google Scholar] [CrossRef]
- Ghosh, U.K.; Islam, M.N.; Siddiqui, M.N.; Khan, M.A.R. Understanding the roles of osmolytes for acclimatizing plants to changing environment: A review of potential mechanism. Plant Signal. Behav. 2021, 16, 1913306. [Google Scholar] [CrossRef]
- Kusale, S.P.; Attar, Y.C.; Sayyed, R.Z.; El Enshasy, H.; Hanapi, S.Z.; Ilyas, N.; Elgorban, A.M.; Bahkali, A.H.; Marraiki, N. Inoculation of Klebsiella variicola alleviated salt stress and improved growth and nutrients in Wheat and Maize. Agronomy 2021, 11, 927. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of proline under changing environments: A review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef]
- Montilla-Bascón, G.; Rubiales, D.; Hebelstrup, K.H.; Mandon, J.; Harren, F.J.M.; Cristescu, S.M.; Mur, L.A.J.; Prats, E. Reduced nitric oxide levels during drought stress promote drought tolerance in barley and is associated with elevated polyamine biosynthesis. Sci. Rep. 2017, 7, 13311. [Google Scholar] [CrossRef]
- Sperdouli, I.; Moustakas, M. Interaction of proline, sugars, and anthocyanins during photosynthetic acclimation of Arabidopsisthaliana to drought stress. J. Plant Physiol. 2012, 169, 577–585. [Google Scholar] [CrossRef]
- Li, H.; Guo, Q.; Jing, Y.; Liu, Z.; Zheng, Z.; Sun, Y.; Xue, Q.; Lai, H. Application of Streptomyces pactum Act12 enhances drought resistance in wheat. J. Plant Growth Regul. 2020, 39, 122–132. [Google Scholar] [CrossRef]
- Moreno-Galván, A.E.; Cortés-Patiño, S.; Romero-Perdomo, F.; Uribe-Vélez, D.; Bashan, Y.; Bonilla, R.R. Proline accumulation and glutathione reductase activity induced by drought-tolerant rhizobacteria as potential mechanisms to alleviate drought stress in Guinea grass. Appl. Soil Ecol. 2020, 147, 103367. [Google Scholar] [CrossRef]
- Garg, A.K.; Kim, J.K.; Owens, T.G.; Ranwala, A.P.; Choi, Y.D.; Kochian, L.V.; Wu, R.J. Trehalose accumulation in rice plants confers high tolerance levels to different abiotic stresses. Proc. Natl. Acad. Sci. USA 2002, 99, 15898–15903. [Google Scholar] [CrossRef]
- Rodríguez-Salazar, J.; Suárez, R.; Caballero-Mellado, J.; Iturriaga, G. Trehalose accumulation in Azospirillumbrasilense improves drought tolerance and biomass in maize plants. FEMS Microbiol. Lett. 2009, 296, 52–59. [Google Scholar] [CrossRef]
- Giri, J. Glycine betaine and abiotic stress tolerance in plants. Plant Signal. Behav. 2011, 6, 1746–1751. [Google Scholar] [CrossRef]
- Niu, T.; Zhang, T.; Qiao, Y.; Wen, P.; Zhai, G.; Liu, E.; Al-Bakre, D.A.; Al-Harbi, M.S.; Gao, X.; Yang, X. Glycine betaine mitigates drought stress-induced oxidative damage in pears. PLoS ONE 2021, 16, e0251389. [Google Scholar] [CrossRef]
- Zhang, H.; Murzello, C.; Sun, Y.; Kim, M.S.; Xie, X.; Jeter, R.M.; Zak, J.C.; Dowd, S.E.; Paré, P.W. Choline and osmotic-stress tolerance induced in Arabidopsis by the soil microbe Bacillus subtilis (GB03). Mol. Plant-Microbe Interact. 2010, 23, 1097–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gou, W.; Zheng, P.; Chen, F.; Zhang, L.; Cui, Z.; Cao, M.; Zhang, L.; Hu, J. Accumulation of choline and glycinebetaine and drought stress tolerance induced in maize (Zea mays) by three plant growth promoting rhizobacteria (PGPR) strains. Pak. J. Bot. 2015, 47, 581–586. [Google Scholar]
- Gowtham, H.G.; Hariprasad, P.; Chandra Nayaka, S.; Niranjana, S.R. Application of rhizobacteria antagonistic to Fusarium oxysporum sp. lycopersici for the management of Fusarium wilt in tomato. Rhizosphere 2016, 2, 72–74. [Google Scholar] [CrossRef]
- Zhou, C.; Ma, Z.; Zhu, L.; Xiao, X.; Xie, Y.; Zhu, J.; Wang, J. Rhizobacterial strain Bacillus megaterium BOFC15 induces cellular polyamine changes that improve plant growth and drought resistance. Int. J. Mol. Sci. 2016, 17, 976. [Google Scholar] [CrossRef]
- Cassán, F.; Maiale, S.; Masciarelli, O.; Vidal, A.; Luna, V.; Ruiz, O. Cadaverine production by Azospirillumbrasilense and its possible role in plant growth promotion and osmotic stress mitigation. Eur. J. Soil Biol. 2009, 45, 12–19. [Google Scholar] [CrossRef]
- Tiwari, S.; Lata, C.; Chauhan, P.S.; Nautiyal, C.S. Pseudomonas putida attunes morphophysiological, biochemical and molecular responses in Cicer arietinum L. during drought stress and recovery. Plant Physiol. Biochem. 2016, 99, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Sagar, A.; Rai, S.; Ilyas, N.; Sayyed, R.Z.; Al-Turki, A.I.; El Enshasy, H.A.; Simarmata, T. Halotolerant rhizobacteria for salinity-stress mitigation: Diversity, mechanisms and molecular approaches. Sustainability 2022, 14, 490. [Google Scholar] [CrossRef]
- Ibort, P.; Molina, S.; Ruiz-Lozano, J.M.; Aroca, R. Molecular insights into the involvement of a never ripe receptor in the interaction between two beneficial soil bacteria and tomato plants under well-watered and drought conditions. Mol. Plant-Microbe Interact. 2018, 31, 633–650. [Google Scholar] [CrossRef]
- Omar, S.A.; Fetyan, N.A.H.; Eldenary, M.E.; Abdelfattah, M.H.; Abd-Elhalim, H.M.; Wrobel, J.; Kalaji, H.M. Alteration in expression level of some growth and stress-related genes after rhizobacteria inoculation to alleviate drought tolerance in sensitive rice genotype. Chem. Biol. Technol. Agric. 2021, 8, 41. [Google Scholar] [CrossRef]
- Sarma, R.K.; Saikia, R. Alleviation of drought stress in mung bean by strain Pseudomonas aeruginosa GGRJ21. Plant Soil 2014, 377, 111–126. [Google Scholar] [CrossRef]
- Hakim, S.; Naqqash, T.; Nawaz, M.S.; Laraib, I.; Siddique, M.J.; Zia, R.; Mirza, M.S.; Imran, A. Rhizosphere engineering with plant growth-promoting microorganisms for agriculture and ecological sustainability. Front. Sustain. Food Syst. 2021, 5, 617157. [Google Scholar] [CrossRef]
- Manasa, M.; Ravinder, P.; Gopalakrishnan, S.; Srinivas, V.; Sayyed, R.Z.; El Enshasy, H.A.; Yahayu, M.; Kee Zuan, A.T.; Kassem, H.S.; Hameeda, B. Co-inoculation of Bacillus spp. for growth promotion and iron fortification in Sorghum. Sustainability 2021, 13, 12091. [Google Scholar] [CrossRef]
- Jabborova, D.; Kannepalli, A.; Davranov, K.; Narimanov, A.; Enakiev, Y.; Syed, A.; Elgorban, A.M.; Bahkali, A.H.; Wirth, S.; Sayyed, R.Z.; et al. Co-inoculation of rhizobacteria promotes growth, yield, and nutrient contents in soybean and improves soil enzymes and nutrients under drought conditions. Sci. Rep. 2021, 11, 22081. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, R.; Nakkeeran, S.; Saranya, N.; Senthilraja, C.; Renukadevi, P.; Krishnamoorthy, A.S.; El Enshasy, H.A.; El-Adawi, H.; Malathi, V.G.; Salmen, S.H.; et al. Mining the genome of Bacillus velezensis VB7 (CP047587) for MAMP genes and non-ribosomal peptide synthetase gene clusters conferring antiviral and antifungal activity. Microorganisms 2021, 9, 2511. [Google Scholar] [CrossRef]
- Khairnar, M.; Hagir, A.; Parmar, K.; Sayyed, R.Z.; James., E.K.; Rahi, P. Phylogenetic diversity and plant growth-promoting activities of rhizobia nodulating fenugreek (Trigonella foenum-graecum Linn.) cultivated in different agroclimatic regions of India. FEMS Microbiol. Ecol. 2022, 98, fiac014. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PGPR Strains | Plant Species | Mechanisms | Beneficial Effects | Reference |
|---|---|---|---|---|
| Bacillus megaterium and Pseudomonas putida | Trifolium repens | IAA production | Enhanced the water content, root and shoot biomass | [24] |
| Bacillus amyloliquefaciens, B. muralis, B. pumilus, B. simplex, B. thuringiensis, Enterobacter aerogenes, Moraxella pluranimalium, and P. stutzeri | T. aestivum | IAA production | Significantly improved the shoot length, spike length, seed weight, tillers and number of spikelets, and increased the peroxidase, acid phosphatase and proline content in plants | [46] |
| Bacillus sp. and Enterobacter sp. | T. aestivum and Z. mays | IAA and salicylic acid production | Displayed the root system architecture alteration viz., increased the number of root tips, root surface area, root length and root branching | [26] |
| P. aeruginosa | Vigna radiata | IAA production | Enhanced the shoot length, number of grains, pod/plant, total yield, 100 seed weight and 100 seed straw weight, improved the photosynthetic activity, membrane stability, relative water content and antioxidant efficacy | [47] |
| B. subtilis | Lactuca sativa | Cytokinin production | Increased the shoot cytokinins, stimulated the shoot mass accumulation and shortened roots, decreased stomatal conductance and root/shoot ratios | [48] |
| B. subtilis | Platycladus orientalis | Cytokinin production | Showed the higher leaf relative water content and water potential, increased the plant root exudates (like amino acids, sugars and organic acids) and stomatal conductance, and elevated the levels of cytokinins in shoot | [49] |
| A. calcoaceticus | S. williamsii | Cytokinin production | Increased the photosynthetic rate, decreased the stomatal conductance and intracellular CO2 concentration | [50] |
| P. putida | Glycine max | Gibberellin production | Increased the shoot length and fresh weight, higher chlorophyll content, lower levels of ABA and salicylic acid, higher jasmonic acid level, reduced sodium content, increased phosphate content and modulated the antioxidants by decreasing radical scavenging activity, SOD and flavonoids | [51] |
| B. licheniformis and P. fluorescens | Vitis vinifera | ABA production | Increased the plant ABA levels, diminished the water loss rate and incremented the synthesis of defense-related terpenes | [52] |
| B. marisflavi | Brassica juncea | ABA analogue/xanthoxin production | Delayed the drooping points of plants and higher drought stress tolerance index, induced the stomatal closure, inhibited the seed germination, and decreased the gibberellic acid induced α-amylase activity | [35] |
| PGPR Strains | Plant Species | Mechanisms | Beneficial Effects | Reference |
|---|---|---|---|---|
| B. subtilis | Trigonella foenum-graecum | ACC deaminase | Improved the plant growth and nutrient uptake, reduced the ACC level, alleviated the ethylene induced damage, increased the rhizobial nodulation and arbuscular mycorrhizal fungal colonization in plants | [82] |
| Achromobacter xylosoxidans, P. oryzihabitans and V. paradoxus | Solanum tuberosum | ACC deaminase | Increased the plant root biomass and tuber yield, decreased the concentrations of rhizosphere ACC and proteinogenic amino acids exuded from the plant root | [83] |
| P. putida | Z. mays | ACC deaminase | Enhanced the growth of seedlings, root colonization and improved the stomatal conductance and cellular metabolites | [84] |
| Burkholderia sp. and Mitsuaria sp. | Arabidopsis thaliana and Z. mays | ACC deaminase and exopolysaccharide production | Altered the root structure system, reduced the evapotranspiration, and modified the level of proline, MDA, phytohormones, and activity of antioxidant enzymes | [25] |
| P. fluorescens and P. palleroniana | T. aestivum | ACC deaminase | Enhanced the plant growth, foliar nutrient content, chlorophyll content, proline content and activity of antioxidant enzymes (APX, CAT, GPX, and SOD), and decreased MDA and H2O2 content | [85] |
| Enterobacter hormaechei, P. fluorescens, and P. migulae | Setaria italica | ACC deaminase and exopolysaccharides production | Stimulated the seed germination and seedling growth, efficiently colonized the root adhering soil and enhanced the root adhering soil/ root tissue ratio and soil moisture content | [70] |
| Consortia of B. subtilis, Ochrobactrum pseudogrignonense, and Pseudomonas sp. | Pisum sativum and Vigna mungo | ACC deaminase | Significantly enhanced the seed germination, dry weight, shoot length and root length, higher leaf chlorophyll content, improved the root recovery intension and relative water content, decreased the ACC accumulation and elevated the production of ROS scavenging antioxidant enzymes and cellular osmolytes | [79] |
| Bacillus sp. and Enterobacter sp. | Mucuna pruriens | ACC deaminase | Significantly improved the root and shoot lengths, reduced the ACC content and ethylene emission, and higher emission of isoprene | [12] |
| Ochrobactrum anthropi, P. fluorescens, P. palleroniana and Variovorax paradoxus | T. aestivum | ACC deaminase | Increased the shoot growth, root growth, root/ shoot ratio, total chlorophyll content, proline content and total phenolics, exhibited lower H2O2 content, reduced MDA levels and improved the activity of antioxidant enzymes (APX, CAT, GPX, and SOD) and foliar nutrient contents (calcium, nitrogen, potassium, phosphorus, and sodium) | [80] |
| B. subtilis and B. thuringiensis | H. annuus | ACC deaminase | Showed the plant growth promotion, increased the proline content and activity of antioxidant enzymes (APX and SOD) and decreased MDA content | [86] |
| O. anthropi, P. fluorescens, P. palleroniana and V. paradoxus | Eleusine coracana | ACC deaminase | Improved the overall plant growth parameters and concentration of nutrients, elevated the activity of ROS scavenging antioxidant enzymes (APX, CAT, GPX, and SOD), cellular osmolytes (such as proline and phenol), higher leaf chlorophyll and reduced the level of H2O2 and MDA | [87] |
| B. subtilis | Solanum lycopersicum | ACC deaminase | Improved the plant growth, relative water content, the activity of antioxidant enzymes (APX and SOD) and proline content, and decreased MDA, H2O2, and superoxide anion accumulation | [78] |
| Bacillus velezensis | Z. mays | ACC deaminase and exopolysaccharides production | Significantly showed better root colonization, improved the plant growth and physiological parameters viz., water use efficiency, vapor pressure, stomatal conductance, photosynthesis, and transpiration | [37] |
| P. fluorescens | Z. mays | ACC deaminase | Significantly improved the concentration of photosynthetic pigments, Fv/Fm ratio, free proline, total soluble sugars and nutrients uptake, and increased the yield traits | [88] |
| P. lini and Serratia plymuthica | Ziziphus jujuba | ACC deaminase | Increased the plant height, root and shoot dry matter, relative water content and soil aggregate stability, decreased the levels of MDA and ABA, and increased the activity of antioxidant enzymes (POD and SOD) | [89] |
| Bacillus sp. and Enterobacter sp. | M. pruriens | ACC deaminase | Showed the higher water use efficiency and total biomass, lower root ACC content and ethylene emission, higher levels of isoprene emission and carbon assimilation | [90] |
| Enterobacter soli and P. corrugata | V. vinifera | ACC deaminase | Increased the soil aggregate stability, root adhering soil/root tissue ratio, levels of phosphorus and nitrogen in plant leaves and soil, altered the biomass of root and shoot, plant height, relative water content and net photosynthetic rate, and changed the contents of IAA, ABA, and MDA in plants | [36] |
| B. amyloliquefaciens | Pennisetum glaucum | ACC deaminase | Increased the seed germination, seedling vigor index, plant growth parameters, total chlorophyll content, relative water content, proline content and activity of antioxidant enzymes (APX and SOD), and decreased the MDA content | [81] |
| P. putida | H. annuus | Exopolysaccharides production | Proficiently colonized the root adhering soil and rhizoplane, enhanced the plant biomass, survival rate, root adhering soil/root tissue ratio and percentage of stable soil aggregates | [91] |
| B. amyloliquefaciens, B. licheniformis, B. subtilis, B. thuringiensis, and Paenibacillus favisporus | Z. mays | Exopolysaccharides production | Increased plant biomass, soil aggregate stability, root adhering soil/root tissue ratio, leaf water potential and relative water content, decreased leaf water loss, enhanced the proline, free amino acids and sugars and decreased the activity of antioxidant enzymes (APX, CAT and GPX) and electrolyte leakage | [71] |
| Alcaligenes faecalis, Proteus penneri, and P. aeruginosa | Z. mays | Exopolysaccharides production | Improved the plant biomass, shoot and root length, soil moisture content, leaf area, relative water content, protein and sugar content, decreased the activity of antioxidant enzymes (CAT, POD and SOD) and proline content | [72] |
| B. endophyticus and P. aeruginosa | A. thaliana | Exopolysaccharides production | Better root colonization and increased plant water content, fresh and dry weights | [73] |
| B. cereus and Planomicrobium chinense | T. aestivum | Exopolysaccharides production | Significantly increased the contents of leaf sugar and protein, higher chlorophyll fluorescence (Fv/Fm), chlorophyll content and performance index, and reduced the activity of antioxidant enzymes (APOX, CAT, and POD), proline content and lipid peroxidation, enhanced the relative water content and augmented the accumulation of micro/ macronutrients (such as Ca, Mg, Na, K, Cu, Cr, Zn, and Fe) | [33] |
| P. azotoformans | T. aestivum | Exopolysaccharides (biofilm) production | Significantly enhanced the root colonization, plant growth characteristics, physiological attributes and photosynthetic pigment efficiency, and decreased the activity of antioxidant enzymes (CAT, GR, and SOD) | [68] |
| P. chlororaphis | A. thaliana | Production of bacterial volatile 2R,3R-butanediol | Reduced the loss of water via stomatal closure | [92] |
| Bacillus thuringiensis | T. aestivum | Reduction in volatile emissions | Enhanced the biomass production, greater photosynthesis and survival of drought stress affected plants | [38] |
| PGPR Strains | Plant Species | Mechanisms | Beneficial Effects | Reference |
|---|---|---|---|---|
| P. aeruginosa | V. radiata | – | Strongly upregulated the drought stress responsive genes such as catalase (CAT1), dehydrin (DHN) and dehydration-responsive element binding protein (DREB2A) | [143] |
| P. putida | Cicer arietinum | Molecular responses | Repressed the expression of transcription activation genes (DREB1A and NAC1) and stress responsive genes (DHN and LEA), upregulated the antioxidative enzymes (APX, CAT, and GST), downregulated the ethylene biosynthesis genes (ACS and ACO), upregulated jasmonate signaling gene (MYC2) and salicylic acid signaling gene (PR1) | [139] |
| B. megaterium and Enterobacter sp. | S. lycopersicum | Molecular responses | Reduced the expression of ethylene biosynthesis genes (ACO, ACS, ERF, ETR, and TCTR1) | [141] |
| Consortia of B. subtilis, O. pseudogrignonense, and Pseudomonas sp. | P. sativum and V. mungo | ACC deaminase | Downregulated ACO gene expression | [79] |
| B. subtilis | S. lycopersicum | ACC deaminase | Significantly decreased the drought responsive gene (Le25) and ethylene responsive factor (SlERF84) | [78] |
| S. pactum | T. aestivum | – | Upregulated the levels of stress resistance gene expression (such as EXPA2, EXPA6, P5CS, and SnRK2) | [128] |
| B. amyloliquefaciens | P. glaucum | ACC deaminase | Significantly increased the expression of antioxidant enzymes (APX1 and SOD1), and decreased the expression of ethylene-responsive factor (ERF-1B) and drought-responsive gene (DREB-1E) | [81] |
| B. megaterium, P. azotoformans and Rhizobium sp. | O. sativa | Molecular responses | Altered the expression of growth and stress related genes (AP2-EREBP, COX1, DHN, EXP1, EXP2, EXP3, GRAM, GST, NAM, and NRAMP6) | [142] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gowtham, H.G.; Singh, S.B.; Shilpa, N.; Aiyaz, M.; Nataraj, K.; Udayashankar, A.C.; Amruthesh, K.N.; Murali, M.; Poczai, P.; Gafur, A.; et al. Insight into Recent Progress and Perspectives in Improvement of Antioxidant Machinery upon PGPR Augmentation in Plants under Drought Stress: A Review. Antioxidants 2022, 11, 1763. https://doi.org/10.3390/antiox11091763
Gowtham HG, Singh SB, Shilpa N, Aiyaz M, Nataraj K, Udayashankar AC, Amruthesh KN, Murali M, Poczai P, Gafur A, et al. Insight into Recent Progress and Perspectives in Improvement of Antioxidant Machinery upon PGPR Augmentation in Plants under Drought Stress: A Review. Antioxidants. 2022; 11(9):1763. https://doi.org/10.3390/antiox11091763
Chicago/Turabian StyleGowtham, Hittanahallikoppal Gajendramurthy, Sudarshana Brijesh Singh, Natarajamurthy Shilpa, Mohammed Aiyaz, Kalegowda Nataraj, Arakere Chunchegowda Udayashankar, Kestur Nagaraj Amruthesh, Mahadevamurthy Murali, Peter Poczai, Abdul Gafur, and et al. 2022. "Insight into Recent Progress and Perspectives in Improvement of Antioxidant Machinery upon PGPR Augmentation in Plants under Drought Stress: A Review" Antioxidants 11, no. 9: 1763. https://doi.org/10.3390/antiox11091763