
Bacillus for Plant Growth Promotion and Stress Resilience: What Have We Learned?

1
Department of Biochemistry, University of Johannesburg, Auckland Park, Johannesburg 2006, South Africa
2
International R&D Division, Omnia Nutriology, Omnia Group (Pty) Ltd., Johannesburg 2021, South Africa
*
Authors to whom correspondence should be addressed.
Plants 2022, 11(19), 2482; https://doi.org/10.3390/plants11192482
Submission received: 3 September 2022
/
Revised: 15 September 2022
/
Accepted: 19 September 2022
/
Published: 22 September 2022
(This article belongs to the Section Plant Protection and Biotic Interactions)
Abstract
:The rhizosphere is a thin film of soil that surrounds plant roots and the primary location of nutrient uptake, and is where important physiological, chemical, and biological activities are occurring. Many microbes invade the rhizosphere and have the capacity to promote plant growth and health. Bacillus spp. is the most prominent plant growth promoting rhizobacteria due to its ability to form long-lived, stress-tolerant spores. Bacillus-plant interactions are driven by chemical languages constructed by a wide spectrum of metabolites and lead to enhanced plant growth and defenses. Thus, this review is a synthesis and a critical assessment of the current literature on the application of Bacillus spp. in agriculture, highlighting gaps that remain to be explored to improve and expand on the Bacillus-based biostimulants. Furthermore, we suggest that omics sciences, with a focus on metabolomics, offer unique opportunities to illuminate the chemical intercommunications between Bacillus and plants, to elucidate biochemical and molecular details on modes of action of Bacillus-based formulations, to generate more actionable insights on cellular and molecular events that explain the Bacillus-induced growth promotion and stress resilience in plants.
1. Introduction
Edaphic factors and genetics play a pivotal role in the growth and yield of crop plants [1]. Over the past decades, genetic engineering and plant breeding approaches have been employed to develop new cultivars with desired traits, such as high yield and resistance to environmental stresses [2]. However, there is a less commercial success for genetically modified crops due to ethical constraints concerning genetically modified organisms (GMO) [3]. To obtain better crop yield, applications of chemical fertilizers have been the opted strategy. However, over time, studies and empirical evidence have shown that this traditional method—the use of chemical fertilizers—is not sustainable due to the inherent negative effects these products have on the environment. The excessive utilization of chemical fertilizers has shown to lead to toxic build-up of heavy metals, soil acidification and soil crust, thereby reducing the soil content of organic matter and humic substance. Soil acidification reduces crop phosphate intake, raises the concentration of harmful ions in the soil, and inhibits crop growth [4].
The incorporation of biostimulants, such as plant growth-promoting rhizobacteria (PGPR)-based formulations, in cropping systems has increasingly shown to be a promising strategy for sustainable agriculture and global food security, aligning with the United Nations sustainable development goals (SDGs) [5]. A broad array of bacterial species has been reported to possess plant growth-promoting attributes with the prominent species belonging to the genus Bacillus. Members of the genus Bacillus are ubiquitous, Gram-positive, and aerobic bacteria [6,7]. Bacillus species produce a multitude of enzymes, antibiotics, and metabolites which give them prominent applications in various sectors such as pharmaceuticals and agriculture. Furthermore, their uniqueness and popularity arise from their spore forming ability which enables them to grow in unconducive environmental conditions [8]. When applied to the plant as dormant cells, these Bacillus spores must germinate to form metabolically active cells. Following germination, these bacteria could be attracted by chemotaxis and leading to the root colonization process (which is mechanistically complex) and exerting growth promotion potentials [9]. Upon colonization, Bacillus spp. elicit direct (e.g., siderophore production, nitrogen fixation, phytohormone production and nutrient solubilization) and indirect mechanisms (such as production of exo-polysaccharides (EPS), biofilm formation, hydrogen cyanide (HCN) and lytic enzymes) to promote plant growth and yield, under various environmental conditions [7].
Thus, this review is a synthesis and a critical assessment of the current literature on the application of Bacillus spp. in agriculture, highlighting gaps that remain to be explored to improve and expand on the Bacillus-based biostimulants that are currently on the market. Considering ongoing efforts to understand the chemical intercommunication between rhizobacteria and plants, we herein review the current knowledgebase on key metabolites secreted by both the bacteria and the plant, their role in the communication between the two organisms, and responses to various environmental stresses. We acknowledge the growing literature and reports on emerging studies and insights on molecular mechanisms underlying rhizobacteria–plant interactions. The intent of this review is to provide a synthesis on the belowground chemical lexicon used for these interactions, with a focus on Bacillus, briefly summarizing the current state of knowledge in this regard. Furthermore, we discuss and emphasize that omics sciences, with a focus on metabolomics, offer unique opportunities to decode the “dark matter” in the chemical intercommunications between Bacillus and plants, to elucidate biochemical and molecular details on modes of action of Bacillus-based formulations, to generate more actionable insights on cellular and molecular events that explain the Bacillus-induced growth promotion and stress resilience in plants.
2. Bacillus spp.-Based Biostimulants and Growth Promotion Mechanisms
Various plant-associated Bacillus spp. have been commercialized as biostimulants for plant protection and growth promotion [5]; and examples of these formulations are given in Table 1. However, there has been a slow rate of exploitation of these Bacillus-based formulations. This is mainly due to a lack of understanding of the chemistry and biochemical mechanisms underlying the modes of action of the Bacillus spp. strains and defining the efficacy of these formulations when applied to crop plants under field conditions. The latter are multifactorial, impacting the growth and productivity of the plant. Various studies have illustrated discrepancies in the performance of rhizobacteria applied under well-controlled conditions and field conditions [10]. There is indeed a need for fundamental studies to understand the metabolism and the biochemistry of Bacillus strains, to elucidate biochemical and molecular events that define Bacillus-plant interactions at different levels and in both systems, i.e., bacteria and host plants.
Despite the lack of fundamental understanding of molecular mechanisms that govern modes of action of Bacillus spp., the applications of Bacillus-based formulations on plants (Table 1) have shown positive impacts such as plant growth promotion. The latter is thought to be due to Bacillus-enhanced nutrient uptake and hormonal modulations. Nutrient availability plays a pivotal role in seed germination and plant growth. However, the bioavailable forms of nutrients such as phosphorus and nitrogen are limited in the rhizosphere. Thus, Bacillus spp. assists in converting the complex forms of these essential nutrients to simple available forms [2]. Bacillus spp. can facilitate the uptake of phosphates by the roots through the secretion of phosphatases and low molecular weight organic acids such as formic acid, acetic acid, lactic acid, glycolic acid, fumaric acid, and succinic acid which acidify the environment to aid the conversion of inorganic phosphates into free phosphate [22]. In addition, Bacillus spp. also produces siderophores which help to solubilize iron from minerals and organic compounds in the rhizospheres. Low molecular weight siderophores such as enterobactin, pyochelin, alcaligin, and rhizoferin have iron-chelating ability making it very arduous for other microbes to access iron. The siderophore–iron complexes can be readily absorbed by the plants; however the transport systems involved therein are not fully understood [23]. Bacillus also facilitates iron mobility by inducing the upregulation of iron acquisition genes in plants. Additionally, studies have shown that Bacillus can increase the concentration of metal ions, which are often a limiting factor for plant growth by breaking them down into nanoparticles thus facilitating their mobilization as illustrated in Figure 1 [24].
Furthermore, upon colonization, Bacillus can directly enhance plant growth by the secretion of cytokine hormones and volatile organic compounds (VOCs) that modify plant hormone networks, promoting cell division and growth [25]. Two VOCs, 3-hydroxy-2-butanone and 2,3-butanediol, which are produced by B. subtilis, have shown to boost plant growth by altering cytokinin and ethylene homeostasis [26]. In another study by Zhang et al. [27], B. subtilis modulated auxin homeostasis by lowering its levels in leaves and inducing optimal growth in the Arabidopsis plant. Spermidine, which is also produced by various Bacillus spp., was reported to promote plant growth via the induction of expansins, and the reduction of ethylene levels in plants [28]. Despite the proposed models that explain these plant growth promotion events induced by Bacillus spp., the chemical language that dictates, mechanistically, Bacillus-plant interactions is still poorly understood. Thus, in the following section, we look at the growing literature on chemical communications between plants and PGPR, pointing out key postulations.
3. Chemical Conversation between Plant and Bacillus spp. Which Leads to Plant Growth Promotion
The communication between plant roots and microbes in the rhizosphere is highly organized and regulated through a dynamic range of specialized metabolites and exudates which ultimately result in altered gene expression in one or both of the interacting partners [29]. The compounds produced by Bacillus spp. include phytohormones, ACC deaminase, volatile organic compounds, polyamines, lipopeptides, and acyl homoserine lactose. On the other hand, plants produce a range of various low-molecular weight molecules such as phytohormones, organic acids, and flavonoids (Figure 2).
The success of microbial based biostimulants relies on the root colonization ability of the beneficial microbes. As abovementioned, root colonization is a complex process which involves a highly dynamic molecular language (which remains enigmatic) between the microbes and the plant in the rhizosphere. Furthermore, this process is influenced by various parameters such as bacterial communities, root architecture and exudates (which depend on the developmental stages and physiological statuses of the plants), (a)biotic factors, and the physicochemical and biological components of the soil [30]. An elaboration on these belowground interactome associates is beyond the scope of this review; however, it suffices here to highlight that understanding the various factors that define the belowground interactions between the bacteria and plant is essential for better and novel exploration of these beneficial bacteria for agricultural practices.
3.1. The Chemical Lexicon of Bacillus spp.-Phytohormones and Organic Compounds
Bacillus spp. are known to produce a wide range of hormones that act as signal molecules in the rhizosphere and involved in the belowground interaction networks (Figure 2). Some of the phytohormones known to be produced by Bacillus spp. include indole acetic acid (IAA), cytokinins (CKs), gibberellins (GAs), and abscisic acid (ABA) [31,32]. IAA is a potent signaling molecule vital for plant-microbe interactions and directly improve plant growth by elevating plant auxin pool which leads to cell elongation, vascular tissue development, and apical dominance [32]. Recent metabolomics and molecular studies have enabled the identification of genes involved in IAA biosynthesis in B. amyloliquefaciens showing that it can occur through multiple pathways [33]. Among the various IAA synthesis pathways that have been identified, tryptophan has been confirmed to be the main precursor [32]. Furthermore, studies have pointed out that plant roots secrete tryptophan in the rhizosphere (Figure 2) which is utilized by rhizobacteria as a precursor for IAA biosynthesis [34,35]. This typically points to the belowground chemical interactomes that define Bacillus–plant interactions.
Bacillus spp. produce a vast range of cytokinins (CKs) (Figure 2) including zeatin, zeatin riboside, zeatin glycoside, izopentyl adenine, and izopentyl adenosine, which have been reported to be produced by B. cereus, B. megaterium, and B. subtilis [36]. Inoculation of lettuce plants with CK-producing B. subtilis increased shoots biomass [25]. Furthermore, the production of zeatin and zeatin riboside by B. subtilis can elicit the exudation of amino acids from the roots in wheat, subsequently increasing the diversity and quantity of beneficial microbiota in the rhizosphere [37]. Another group of phytohormones that is produced and secreted by Bacillus spp. is gibberellin (Figure 2) which is also involved in different plant developmental processes and the regulation of many physiological processes [38]. In the lettuce plant, the production of gibberellic acid by Bacillus spp. has been associated with an increase in nutritional metabolites such as amino acids, macro-and micronutrients, fructose, and carotenoids, thus increasing the quality of the crop [39].
In addition to hormones, rhizobacteria are reported to use volatile organic compounds (VOCs) in the chemical interactions with plants or other microorganisms (Table 2). Studies have shown that various genera of bacteria can regulate plant growth from a distance [40]. Some of these VOCs exuded by bacteria act as growth promoter molecules [41]. VOCs are low molecular weight and high vapor pressure compounds and act as signal molecules over short distances [42]. These features enable VOCs to facilitate intercellular and organismal interactions [43]. VOCs from Bacillus spp. are categorized into aldehydes, ketones, alkyls, alcohols, alkenes, esters, acids, ethers, heterocyclic, and phenolic compounds; however, many of these VOCs are still unknown [44,45]. Due to the structural diversity of these microbial VOCs, many natural functions have been inferred as highlighted in Table 2. For example, the production of the VOCs albuterol and 1,3-propanediol by B. subtilis induced differential expression of genes involved in the biosynthesis of the phytohormones auxin, gibberellin, cytokinin, and ethylene [42]. This subsequently leads to altered levels of the endogenous content of the related hormones in the roots and leaves, suggesting the involvement of these hormones in signal transduction pathways induced by VOCs which ultimately enhances plant growth. The natural function of these microbial VOCs is not only limited to plant growth but also infer protection against biotic stresses and induce plant tolerance against abiotic stresses. Investigations of these Bacillus VOCs-mediated effects on plants have focused on how they influence the signaling of phytohormones such as salicylic acid (SA), jasmonic acid (JA), ethylene, auxin, etc. [40]. Even though the effects of these Bacillus VOCs on plants are well documented, the underlying signaling mechanisms are still poorly understood [46].
As abovementioned, the belowground chemical interactome is highly complex and dynamic. One of the factors that define the performance of these PGPR is the efficacy in the root colonization by the bacteria, which is highly influenced by root exudates. The latter depends on the developmental stages and physiology of the plants. Furthermore, the microorganisms in the rhizosphere have diverse roles in supporting plant growth, development, and inhibition of host pathogens. This implies interdependency between the host and microbes in the aboveground and belowground interactions. Thus, understanding rhizosphere colonization mechanisms by PGPR is essential for generating inoculants able to compete and efficiently colonize the rhizosphere of plant crops and having a great impact on crop production and more consistent results.
3.2. Plant Root Exudates in Plant–Bacillus Crosstalk
Root exudates are a complex mixture of primary and secondary metabolites released into the rhizosphere by the roots and they maintain constant communication between plants and microbes in the rhizosphere [52]. Root exudates are composed of amino acids, organic acids, phenolics, sugars, and proteins with their highly diversified chemical properties based on different plant species, plant growth stages and the microbiota in the rhizosphere [53]. One of the roles of root exudates is to serve as signals for root colonization by beneficial microbes [54] as shown in Figure 2. PGPR can colonize the roots through chemotaxis, which is the mobility capacity of the bacteria along a chemical signal gradient [55]. Thus, root exudates are the architects of the rhizosphere and congruently the colonization of bacteria on the roots can modulate the metabolites exudation pattern [56].
The bacterial chemotaxis is elicited when a root exudate molecule binds to its cognate receptor [57]. Given the chemical diversity of root exudates, systematic identification of chemo-attractants and chemo-repellents in root exudates and elucidation of how these various root-secreted compounds are detected by multiple chemoreceptors of a PGPR strain can provide a comprehensive understanding of how PGPR colonizes the rhizosphere. In a study by Feng et al. [57], 39 chemo-attractants and 5 chemo-repellents were identified from cucumber root exudates for a well-studied PGPR strain B. amyloliquefaciens SQR9. In this study a mutant strain with 8 putative chemoreceptors deleted, lost chemotactic response to all the 44 compounds showing the importance of these chemoreceptors in plant–PGPR interactions and subsequent colonization. Further characterization of these chemo-effectors (root exudates) and chemoreceptors will broaden our insights into Bacillus–root interaction and provide valuable information to enhance the rhizosphere colonization ability of Bacillus spp., which will promote their application in agricultural production.
Post root colonization by Bacillus, the bacteria proliferate by receiving key signaling compounds and nutrients from the root exudates which ultimately lead to biofilm formulation on the root system which is an indication of a successful root colonization [58]. A study by Yuan et al. [58] showed that root exudates of banana containing organic acids induced both chemotaxis and biofilm formation in B. amyloliquefaciens. In this study, malic acid showed the greatest chemotactic response whereas fumaric acid significantly induced biofilm formation. Root exudates can be modified by environmental perturbations and thus influence the plant–PGPR interaction as it was illustrated in a study by Cesari et al. [59] where drought conditions modified the pattern of molecules exuded by roots, increasing the exudation of naringenin, oleic, citric, and lactic acids, and stimulating the release of terpenes of known antioxidant and antimicrobial activity. Changes in the molecular profiles of these exudates due to drought allowed for enhanced interaction between the roots and PGPR thus reversing the negative effects of drought condition on plant growth. These findings can assist in the formulation of biostimulant inoculants containing key molecules exuded during stress which can improve plant–PGPR interactions and thus promote plant growth and enhance defenses against various stresses.
4. Bacillus spp. Confers Protection to the Plant from Environmental Stresses
Major yield deficiencies, crop damage, and changes in growth rates of plants are caused by abiotic and biotic stresses. Many strategies have been proposed to combat the negative impacts of environmental stresses, and these have included, for instance, the overexpression of single genes encoding enzymes involved in the transportation of ions and scavenging of reactive oxygen species (ROS) being the most popular over the years. The application of this approach is limited due to the resultant pleiotropic effects on the growth of the plant and multiple pathways involved in response to environmental stress [60]. Another strategy is the use of chemical-based fertilizers and pesticides which are increasingly becoming harmful to the environment and some of these xenobiotics have become highly toxic in the agro-food chain. Therefore, nonharmful, environmentally friendly, sustainable, and nature-inspired solutions and strategies are urgently needed, and these include the use of PGPR formulations. The latter can aid in alleviating the effects of environmental stress. Several studies have demonstrated that Bacillus spp.-based biostimulants trigger plant protective mechanisms against both abiotic and biotic stresses.
4.1. Bacillus spp. against Biotic Stress
Biological control, utilizing beneficial microbes, is an excellent approach to limiting the adverse effect of disease-causing microbes on plant health and productivity. Considerable effort has been placed on identifying microbial biocontrol agents that can repress phytopathogens, especially those that are responsible for soil-borne diseases, and that can enhance agricultural productivity [61,62]. Many strains of Bacillus exhibit the ability to act as biocontrol agents against pathogens and thus can be used to suppress diseases [63]. Several mechanisms, both direct and indirect, are responsible for their ability to control pathogenic microbes as shown in Figure 3. These include the production of a wide array of antibiotic compounds (lipopeptides), the ability to form endospores, the ability to form biofilms on root surfaces, and the ability to induce host systemic host resistance, and stimulate plant growth [64].
Biotic stresses often lead to the production of reactive oxygen species (ROS) which lead to oxidative stress and are toxic to the cells. The inoculation of plants with Bacillus-based formulations has shown to elicit the production of antioxidant defense enzymes such as superoxide dismutase and peroxidase, which scavenge the ROS [65]. In a study by Zebelo et al. [66], cotton plants inoculated with Bacillus spp. demonstrated an increase in gossypol and jasmonic acid levels and secretion reducing larval feeding by spodoptera exigua. There was also up-regulation of genes involved in synthesis of allelochemicals and jasmonates in the inoculated plants. Furthermore, Bacillus spp. secrete various catabolic enzymes such as proteases, chitinases, and glucanases, as well as peptide antibiotics and secondary metabolites that contribute to pathogen suppression [40]. Bacillus spp. secrete cyclic lipopeptides such as iturin and surfactin (Figure 3) which play a role in disease suppression by acting as bi-functional molecules through their antifungal activity and elicitation of induced systemic resistance (ISR). The latter involves pathogen recognition at plant cell surface, stimulation of early cellular immune-related events, systemic signaling through a fine-tuned-hormonal cross talk and activation of defense mechanisms [67]. Bacillus spp. can elicit ISR in plants, which switches on pathogenesis related genes, mediated by phytohormone signaling pathways and defense regulatory proteins to pre-condition plants against future pathogen ambush [68].
Bacillus spp. secrete the secondary metabolites, cyclic lipopeptides (CLPs) which are involved in developmental processes such as motility and biofilm formation as well as biocontrol primarily based on their antimicrobial activity [69]. The synthesis of these CLPs is accomplished by multimodular peptide synthetases and depends on a functional phospho-pantheinyl transferase (Sfp) which transfers 4′-phosphopantetheine from coenzyme A to the carrier proteins during non-ribosomal synthesis [70]. Some CLPs secreted by Bacillus have emerged as plant immunity elicitors (Figure 3). The best-described Bacillus CLP is surfactin. When applied as pure compound on roots, surfactin induced ISR in bean, tomato, tobacco, against B. cinerea, in melon against Podosphaera fusca, and peanut, against Sclerotium rolfsii [71,72,73,74]. The Bacillus CLP iturin has shown high bioactivity when applied to leaves as compared to the roots. The biochemical and molecular mechanisms that lead to this contrast are not well understood [75].
Bacillus strains also produce volatile organic compounds which act as elicitors of plant immunity. The most characterized VOCs secreted by Bacillus are 2,3-butanediol and acetoin (Figure 3) produced from glucose in the central metabolism [76]. In a study by Rudrappa et al. [77], exogenous application of acetoin triggered ISR and protected the maize plant against Pseudomonas syringae. What is intriguing in this study is that the expression of acetolactate synthase (an enzyme involved in the synthesis of acetoin) was significantly upregulated in the presence of maize root exudates during the late exponential growth phase, suggesting that root exudates play a role in eliciting acetoin biosynthesis in Bacillus [78]. The effects exerted by these secondary metabolites produced by Bacillus on plant health still need further elucidation as the response of the plant in the tandem presence of the beneficial Bacillus and the pathogen is complex.
4.2. Bacillus spp. against Abiotic Stress
Being sessile organisms, plants have to withstand various adverse abiotic stresses such as drought, salinity, heat/or cold, and heavy metal toxicity which pose a major threat to agriculture by negatively impacting plant growth and yield worldwide [79]. These stresses elicit stress responses in plants, including an accumulation of reactive oxygen species (ROS) and reduced photosynthetic activity, which ultimately leads to reduced plant growth and crop yield. PGPR such as Bacillus spp. can mediate the induction of abiotic stress responses in plants [80]. These responses to abiotic stresses are attributed to metabolic regulations which often require wide changes in the concentration, composition, and distribution of both primary and secondary metabolites. Biostimulants containing Bacillus strains have shown the potential to stimulate abiotic stress tolerance [81]. However, the biochemical and molecular mechanisms governing this Bacillus-induced stress resistance and tolerance are still enigmatic. Mechanisms suggested including changes of phytohormone levels through the secretion of phytohormones by Bacillus or ACC deaminase activity that decrease ethylene levels [82].
Cytokinins such as zeatin produced by Bacillus play a pivotal role in maintaining cellular proliferation and differentiation [83]. A study by Hussain and Hasnain [84] showed that the extracts obtained from cell cultures of B. licheniformis and B. subtilis are capable of increasing the weight and size of cucumber cotyledons separated from the seedlings, by inducing plant cell division due to the presence of cytokinins zeatin and zeatin ribose thus enhancing tolerance to salt stress. Cytokinin production has also been related to the ability of Bacillus species, such as B. subtilis, to increase plant tolerance to drought in lettuce [85] and Platycladus orientalis [86], or against salinity in wheat, due to the production of Zeatin, promoting plant growth [87]. Bacillus also produces auxins such as indole-acetic acid (IAA) which has been reported to confer tolerance against heavy metals [87,88]. For example, the study by Sun et al. [88] demonstrated that the application of IAA-producing Bacillus altitudinis alleviates iron stress in Triticum aestivum L. seedlings by both bioleaching of iron and up-regulation of genes encoding ferritins. Another study by Ji et al. [87] also showed that wheat treatment with IAA-producing B. subtilis can increase plant tolerance under salinity conditions.
Bacillus is also capable of producing gibberellic acids (GAs) which is a group of phytohormones involved in the processes of seed germination, flower initiation, leaf expansion, stem elongation, or flower and fruit development [89]. Various Bacillus species have the ability to produce a wide range of different GAs in vitro [90]. In a study by Ji et al. [87], the production of GAs by B. subtilis was related to its ability to increase the tolerance of wheat under salinity. Moreover, a study by Kang et al. [91] showed that the production of various GA compounds by B. tequilensis was involved in the induction of thermotolerance in soybean plants due to an increase of endogenous jasmonic acid and salicylic acid contents and a downregulation of abscisic acid showing the interaction between the various phytohormones in stress regulation. Abscisic acid (ABA) is considered an essential messenger in the adaptive response of plants to abiotic stress. Under water-deficit conditions, ABA plays a vital role in providing plants the ability to signal to their shoots that they are experiencing stressful conditions around the roots, eventually resulting in water-saving anti-transpirant activity, notably stomatal closure and reduced leaf expansion [92]. ABA upregulates the processes involved in cell turgor maintenance and synthesis of osmoprotectants and antioxidant enzymes conferring desiccation tolerance [93]. Study by Zhang et al. [94] reported a proportional increase in ABA concentration upon exposure of plants to salinity. A study by Pan et al. [95] also reported that the production of ABA by B. subtilis resulted in an increased tolerance to the heavy metal cadmium in Brassica chinesis plant by reducing cadmium-induced photosynthesis inhibition and the oxidative damage in plant tissues through increased levels of antioxidant compounds.
Although many studies have shown that there is a positive correlation between Bacillus synthesized phytohormones and plant growth and development under various abiotic stresses, there is a lack of knowledge of the underlying biochemical mechanisms involved in the enhanced responses to abiotic stresses. Thus, to decipher all the chemical spheres and pathways involved in the interaction between Bacillus phytohormones and the plant under various environmental stresses, there is a need for more systemic biology studies. Apart from phytohormones, Bacillus spp. also produce VOCs, which induce abiotic stress tolerance. Bacillus VOCs have the potential to simultaneously up-regulate high affinity HKT1 in shoots and down-regulate it in the roots, thus enhancing HKT1-dependent shoot-to-root sodium ion circulation, which is crucial in salinity tolerance. This was observed in a study by Zhang et al. [96] that VOCs such as 2,3-butanediol produced by B. amyloliquefaciens GB03 strain reduced accumulation of sodium ions in Arabidopsis shoots resulting in enhanced salinity stress tolerance. Zhang et al. [97] reported that exposure of Arabidopsis to Bacillus VOCs resulted in the accumulation of high levels of choline and glycine betaine, which are vital osmo-protectants that confer drought tolerance in plants [98].
The molecular mechanisms governing Bacillus-mediated abiotic stress tolerance are not limited to Bacillus’ ability to produce phytohormones and VOCs. Other mechanisms associated with enhancing stress tolerance involve triggering biological and physiological processes such as ROS detoxification mechanisms, osmoprotection, stomatal regulation, membrane stability, xylem hydraulic conductance, root zone water, and nutrient availability and metal chelation [99,100,101]. For example, recent studies by Nephali et al. [11], Lephatsi et al. [13] and Othibeng et al. [102] revealed that the application of Bacillus consortium (a commercial biostimulant known as Bacstim® 100) to maize plants improves drought tolerance via mechanism such as enhanced energy production, enhanced expression of drought stress responsive defense genes, osmoregulation, redox homeostasis, strengthening of the plant cell wall and membrane remodeling.
As highlighted herein, there is vast knowledge on the Bacillus–plant interactions leading to growth promotion and stress resistance. However, there is still a lack of fundamental research that provides direct mechanisms of action underlying Bacillus–plant interactions. Thus, further research on the components and effects of Bacillus-based biostimulants on the plant health and growth is required to advance the development of a scientifically based biostimulant industry with the potential to encourage effective exploration and application of Bacillus-based biostimulants in agriculture for improved and sustainable food security. Comprehensive study approaches such as “omics” sciences have shown great potential to interrogate the metabolism of plants and Bacillus to generate mechanistic models with actionable knowledge and insights on how Bacillus promotes plant growth and enhances defense responses.
5. Omics Sciences in Studying Bacillus spp. and Its Effects on Plants: Metabolomics Applications
Omics refers to a biological study field where a comprehensive assessment of molecular entities is performed at different levels in the systems biology, such as DNA (genomics), RNA (transcriptomics), proteins (proteomics), and other cellular molecules (metabolomics) ([103]; Figure 4). Advancements in these technologies have proven instrumental in decoding important multilayered biochemical events underpinning the effects of Bacillus-based biostimulants on plants [99,104,105,106]. Metabolomics is a multidisciplinary omics science which offers a comprehensive identification and quantitation of low-molecular-weight molecules, namely metabolites. Plant metabolites are considered the “cause of plant phenotype” [107]. As such, the application of metabolomics in investigating the rhizospheric complex and dynamic chemical communications between Bacillus and plants would be of great value to agricultural research [108].
The belowground intercommunication involves inter-and intra-organismal interactions (i.e., plant–Bacillus, plant-to-plant, and Bacillus–other microbes) [109]. Such multidimensional interactions are facilitated and controlled through the interconnected belowground signaling networks, resulting in altered plant growth and development. Thus far, as highlighted in Section 3, the known signaling molecules and metabolites include root exudates, volatile organic molecules (VOCs), allelochemicals, lytic enzymes, toxins, and quorum sensing molecules (QSM), phytohormones, siderophores, lipopeptides, and lytic enzymes [110,111]. While some studies have made significant contributions to characterizing the composition of rhizobiomes, there are still considerable gaps in understanding how plants shape their rhizobiomes and how the bacterial presence and exudates affect the plant and its root microbiota [29,112].
Considering the intricate complexities in the rhizosphere mentioned above, studying these interactions concertedly is challenging and almost impractical. Thus, the existing literature mainly focuses on investigating one or two of the interacting partners to intently extract useful and accurate information that can offer the best description of these dynamic interactions. As such, the following sections discuss metabolomics studies aimed at investigating (i) Bacillus intracellular and extracellular chemical space, (ii) interactions between Bacillus and other microbes in the rhizosphere, and (iii) Bacillus and plant intercommunications.
5.1. Application of Metabolomics in Investigating Bacillus Chemical Space
The emergence of omics sciences such as metabolomics allows a comprehensive examination of biological systems at a global level and the exploration of interrelationships and dynamics between the multi-component systems, i.e., holobiont [113]. The untargeted metabolomics studies interrogating the chemical space of Bacillus are still very limited; however, most studies employ analytical platforms such as chromatography coupled to mass spectrometry and nuclear magnetic resonance (NMR) spectroscopy (commonly utilized in metabolomics) to structurally characterize bacterial metabolites (Table 3). For example, the application of LC-MS and NMR spectroscopy revealed the identity of two novel cyclic depsipeptides turnagainolides A and B and that these compounds contain a combination of R and S amino acids and the (E)-3- hydroxy-5-phenylpent-4-enoic acid fragment Hppa, which are hallmarks of nonribosomal peptide biosynthesis [114].
In another study, LC-MS and NMR spectroscopy-guided metabolic profiling was employed and two new cyclic-lipotetrapeptides, bacilotetrins A and B were identified in the extracellular milieu of B. subtilis [115]. A cyclic lipopeptide plipastatin A1 was identified from the extracellular milieu B. amyloliquefaciens SH-B74 using a combination of analytical platforms including LC-MS, GC-MS, and NMR [119]. The cumulative findings of such studies led to wide-ranging secondary metabolites that are known to be present in various Bacillus strains (Table 3). Such metabolites include peptides of low molecular weight that are generated ribosomally (bacteriocins) or non-ribosomally; lipopeptides such as surfactins, iturins, and fengycins and polyketides such as macrolactins, difficidins, and oxidifficidins [116]. Interrogating and profiling the intracellular and extracellular metabolome of the Bacillus strains will provide insightful science-based information that can be leveraged in designing and formulating novel Bacillus biostimulants. Moreover, understanding the intracellular metabolome of Bacillus and the metabolites they secrete to the exterior, can enhance the understanding of Bacillus-plant intercommunication [12,133].
With innovative developments in analytical technologies (integrating artificial intelligence, AI and machine learning, ML), advancements in chemometrics and statistical methods (big data analytics and management), and the integration of orthogonal biological approaches, untargeted metabolomics is becoming a fundamental fulcrum to creating a “Rosetta stone” for deciphering the encryptions existing in the rhizosphere ecology [134,135]. One of the emerging analytical aspects is ion mobility technologies such as ion mobility spectrometry (IMS). Interfacing IMS with mass spectrometry (IMS-MS) increases the analytical power, enabling the efficient separation, resolution, identification, and multidimensional structural characterization of analytes [136,137]. Currently, there are limited studies demonstrating the application of IMS technologies in investigating the Bacillus spp. One of the very few studies include a recent study by Ratiu et al. [138], where a portable aspiration-type ion mobility spectrometer (a-IMS) and gas chromatography-mass spectrometry (GC-MS) were applied to discriminate between different bacteria, Escherichia coli, Bacillus subtilis and Staphylococcus aureus, by rapid sensing of the bacterial metabolic volatiles, which produced differential metabolic fingerprints.
Another analytical tool that has proven instrumental for profiling live microbial colonies is nanoDESI MS [134]. The applications of this technique allowed highly sensitive metabolic profiling directly off living microbial communities, requiring zero sample preparation. Other advantages of this tool include the ability to capture a wide variety of metabolite families within a single mass spectrum directly from a live specimen thus giving comprehensive visuals into the chemical space of bacteria. Such insights are fundamental to describing the microbial chemotype that are more accurate to the phenotype [134]. For example, in this study by Watrous et al. [134], nanoDESI MS and MS/MS molecular networks demonstrated an elevation of metabolite content within B. subtilis across the 60 h time growth period, with increased production of structural variants of the cyclic peptides surfactin, plipastatin, and subtilosin steadily increasing over time.
Furthermore, some of the recent advancements in data handling and metabolite annotation include fourth industrial revolution (4IR)-inspired computational tools such as molecular networking (MN) integrated with in silico annotation tools [139,140,141] are positively impacting the chemical and biological interpretation of untargeted metabolomics studies on Bacillus and Bacillus–plant interactions. The application of MN in microbial metabolomics enabled detection and visualization of related metabolites in Bacillus cells via spectral similarities within and between data sets as chemical families are grouped together [12,134]. For instance, the study by Nephali et al. [12] applied MN tools to profile the intracellular chemical space of PGPR Bacillus strains: B. laterosporus, B. amyloliquefaciens, B. licheniformis 1001, and B. licheniformis M017 and their consortium—this study showed higher content of surfactins in Bacillus consortium and B. amyloliquefaciens and higher content of lichenysins B. licheniformis strains compared to other strains under study. Additionally, MN proved instrumental to uncovering the temporal dynamics in B. subtilis (Watrous et al., 2012) and B. laterosporus, B. amyloliquefaciens, B. licheniformis 1001, and B. licheniformis M017 [12]. Time-dependent MN showed an increasing number of lipopeptides over time in B. amyloliquefaciens, B. licheniformis 1001, B. licheniformis M017, and B. laterosporus and their consortium, and the stationary phase showed the highest content of lipopeptides in the cells [12]. A recent study by Wang et al. [142] revealed novel compounds Hetiamacin E and F from B. subtilis PJS using MS/MS-based MN. Another study by Purves et al. [143] demonstrated the application of metabolomics and MN for microbial secondary metabolite bioprospecting. Decoding metabolic profiles of Bacillus strains will contributes toward understanding the chemical space of PGPR, which will subsequently help in decoding the chemical communication underlying PGPR-plant interactions.
Despite the increasing advancements in the metabolomics workflow mentioned above, the chemical space of Bacillus is still not fully elucidated. Currently, the metabolites that have been found in Bacillus include amino acids, hormones, organic acids, nucleotides, cofactors, and sugar-phosphates [144] and structurally diverse secondary metabolites, such as lipopeptides (e.g., iturins, fengycins, and surfactins), polyketides, polypeptides, macrolactones, lipoamides, fatty acids, isocoumarins [53,145,146], siderophores such as hydroxamates (e.g., schizokinen, pyochelin), catecholates (e.g., bacillibactin, petrobactin), and carboxylates (e.g., rhizobactin) [147,148]. Identification of these metabolites in Bacillus cells have contributed toward understanding the biochemistry of Bacillus and their interaction with the host plants and other microorganism. For example, metabolites such as lipopeptides, macrolides, and polyketide have been demonstrated to possess antimicrobial and antifungal activities (Table 2), mechanisms that are involved in biocontrol of phytopathogens [148]. Moreover, other bacterial metabolites such as hormones (indole acetic acid) and amino acids (tryptophan) have been shown to promote plant growth [148]. However, the current knowledge is barely a tip of the iceberg, as a large percentage of Bacillus metabolome remains unexplored [144,149]. Comprehensively annotating and characterizing the metabolome of Bacillus will advance our understanding of belowground chemical communications between Bacillus and plants. Such accurate models would explain mechanisms of action of Bacillus, molecular events that govern Bacillus-mediated enhanced plant growth and stress protection. This actionable knowledge is necessary for innovatively designing and implementing Bacillus-based formulations for sustainable agriculture.
5.2. Application of Metabolomics in Investigating Bacillus and Other Microorganisms’ Interactions in the Rhizosphere
The holobiont dynamics (assembly of the phytomicrobiome) involves the chemical intercommunications between the microbe–microbe interactions and plant–microbe interactions [150]. As rhizosphere-dwelling bacteria, Bacillus are surrounded and constantly interacting with other microbes via different types of communications [151]. The intercellular interactions between microbes occur via four main mechanisms such as cell–cell signaling, production of secondary metabolites, cell–cell contacts and metabolic interplay [152]. The cell–cell signaling involves the interexchange of diverse chemical signaling molecules such as volatiles, quorum sensing signals, and secondary metabolites to communicate, regulate, and synchronise microbial behaviors. However, this signal network is not well understood—there are knowledge gaps in determining the predominance, diversity, and function of signaling molecules and how such factors drive the communal behavior and population dynamics within phytomicrobiome ecosystem.
As such, metabolomics approaches can be employed to bridge the existing knowledge gaps [108,150,153]. One of the very few is a study by Wen et al. [154], which demonstrated that several bacterial groups such as Bacillus and Chitinophaga were negatively related to the pathogen abundance. The GC-MS analyses revealed significantly different metabolomes in two groups of rhizosphere soils, i.e., the rhizosphere soil of lower harbored more sugars such as fructose, sucrose and melibiose than that in high pathogen abundance, indicated their potential biocontrol ability. A recent study by Andric et al. [155] applied a highly advanced analytical platform, MALDI-FT-ICR MS imaging, revealing that Bacillus mobilizes its cyclic lipopeptide surfactin to improve motility and reduce the toxicity of Pseudomonas by acting as a chemical deactivator of Pseudomonas lipopeptides, sessilins and tolaasins.
The secretion of lipopeptides surfactins by Bacillus displays an antagonist relationship with other microbes and has also been shown to exert a symbiotic relationship with other bacterial species, thus shaping the plant microbiome. For example, the study by Luzzatto-Knaan et al. [133], demonstrated the role of surfactins as an interspecies recruitment factor. In this study by Luzzatto-Knaan et al. [133], mass spectrometry and MN approaches were applied and revealed that B. subtilis secretes surfactins to recruit Paenibacillus dendritiformis to its ecological niche and that P. dendritiformis actively degrades surfactins originating from B. subtilis and marks its territory by accumulating the resulting surfactin degradation products. Such studies illustrate the indispensability of metabolomics in revealing the ecological roles of secondary metabolites such as lipopeptides during microbial interspecies interactions as well as the regulation of their expression under naturally competitive soil conditions [133,155]. Moreover, studies like these bring the scientific community much closer to fully understanding the mechanisms of Bacillus mechanisms for agricultural applications.
5.3. Application of Metabolomics in Investigating the Bacillus and Plant Intercommunications
Various metabolomics studies have illustrated that the Bacillus–plant interaction (which may involve the secretion of secondary metabolites from Bacillus, Table 2) can confer enhanced growth-promotion and defense priming of the plant through the reprogramming of the plant metabolome [146]. For example, a study by Nephali et al. [11] revealed an increased pool of tricarboxylic acid (TCA) intermediates, reprogramming of amino acid profiles and differential changes in phenolics and lipids as key metabolomics signatures induced by the application of Bacillus-based biostimulants on maize plants. Another metabolomics by Kang et al. [156], showed decreased levels of glucose, fructose, sucrose, and trehalose in B. simplex-treated soybean roots compared to the control group—decreased sugar levels was translated to reduced food sources for nematodes. Furthermore, treatment with B. simplex led to higher levels of melibiose, gluconic acid, lactic acid, phytosphingosine, and noradrenaline in soybean roots, which promoted nematocidal activity thus, improving disease resistance.
A metabolomics approach was also applied to reveal the underlying mechanisms employed by Pa. alvei NAS-6G6 and B. velezensis N54 in plant protection against biotic (Fusarium pseudograminearum crown rot) and abiotic (drought) stress. In this study, Pa. alvei NAS-6G6 was found to induce unique protection capacity against biotic and abiotic stress, and combined stresses by upregulating different defense metabolites S. bicolor plants and altering metabolic pathways such as riboflavin metabolism under biotic and drought stress and glutathione metabolism under combined biotic (Fusarium pseudograminearum crown rot) and abiotic (drought) stress. B. velezensis N54 upregulated arginine and proline metabolism, Pa. alvei NAS-6G6 upregulated riboflavin metabolism under biotic stress condition (Fusarium pseudograminearum crown rot) [157]. Riboflavin and proline are known to plays significant roles in regulating antioxidant mechanisms and osmoprotection [157]. A recent metabolomics study by Shahid et al. [146], profiled extracellular secondary metabolites and hormones from Bacillus spp., which positively correlated to plant growth promotion and antifungal properties. The application of metabolomics to investigate the effects of Bacillus and Bacillus-based formulations is gaining some momentum, thus generating a wealth of knowledge driving toward the complete elucidation of Bacillus-mediated plant growth promotion and stress protection mechanisms. Thus, a reader is referred to review papers by Nephali et al. [99] and Lephatsi et al. [13] for more details on applying metabolomics approaches in studying microbe-plant–stress interactions.
Bacillus spp. are emerging as key microorganisms in the biostimulant industry for maintaining sustainable food security. However, as reviewed in this manuscript there are still major knowledge gaps and bottlenecks in the formulation of Bacillus-based biostimulants. This includes poor characterization of active components and synergies in the biostimulant composition and undefined mechanisms and modes of action of biostimulant products, at cellular and molecular levels. Moreover, it is difficult to predict the field application efficacy of these biostimulants due to various environmental factors that may affect field crops, such the soil chemistry, biotic and abiotic stresses. These limitations hinder not only the designing of novel formulations and biostimulant-based agricultural strategies but also the establishment of a standardized legislative framework and regulatory system for the biostimulant industry. Metabolomics, a multidisciplinary omics science, offers unique opportunities to predictively decode the mechanisms and modes of action of biostimulants on crop plants, and elucidating signatory markers and metabolic profiles that define the biostimulant action (Table 2). The application of metabolomics in biostimulant research can help us to comprehensively decipher the molecular basis behind biostimulant-induced plant metabolomic reprograming which leads to improved plant health, growth, and increased yield. Such fundamental and indispensable knowledgebase would improve our understanding of biostimulants, also providing a roadmap for translational applications—designing of novel formulations and devising biostimulant-based precision agricultural programs and modules—for sustainable food security.
6. Conclusions and Future Prospects
In this critical review we narratively revisited the importance of PGPR such as those belonging to the genus Bacillus in improving plant growth and enhancing plant protection against adverse environmental factors. Furthermore, the incorporation of biostimulants, such as microbial formulations (e.g., Bacillus-based products), in cropping systems has increasingly shown to be a promising strategy for sustainable agriculture and global food security, aligning with the United Nations sustainable development goals (SDGs). Despite the current wealth of knowledge on Bacillus spp. and its applications, molecular mechanisms that govern modes of action of Bacillus-based formulations remain poorly understood. Thus, this review suggests that omics sciences, with a focus on metabolomics, offer unique opportunities to illuminate the chemical intercommunications between Bacillus and plants, and to elucidate biochemical and molecular details on modes of action of Bacillus-based formulations.
Author Contributions
Conceptualization, T.T. and F.T.; writing—original draft preparation, T.T., M.M. and L.N.; writing—review and editing, F.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Kleinwechter, U.; Gastelo, M.; Ritchie, J.; Nelson, G.; Asseng, S. Simulating cultivar variations in potato yields for contrasting environments. Agric. Syst. 2016, 145, 51–63. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Hashem, A.; Abd Allah, E.F. Bacillus: A biological tool for crop improvement through bio-molecular changes in adverse environments. Front. Physiol. 2017, 8, 667. [Google Scholar] [CrossRef] [PubMed]
- Bawa, A.S.; Anilakumar, K.R. Genetically modified foods: Safety, risks and public concerns—A review. J. Food Sci. Technol. 2013, 50, 1035–1046. [Google Scholar] [CrossRef]
- Alengebawy, A.; Abdelkhalek, S.T.; Qureshi, S.R.; Wang, M.Q. Heavy metals and pesticides toxicity in agricultural soil and plants: Ecological risks and human health implications. Toxics 2021, 9, 42. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant Growth-Promoting Rhizobacteria: Context, Mechanisms of Action, and Roadmap to Commercialization of Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef]
- Grover, M.; Ali, S.Z.; Sandhya, V.; Rasul, A.; Venkateswarlu, B. Role of microorganisms in adaptation of agriculture crops to abiotic stresses. World J. Microbiol. Biotechnol. 2011, 27, 1231–1240. [Google Scholar] [CrossRef]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of plant growth promoting rhizobacteria in agricultural sustainability—A review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Siddikee, M.A.; Chauhan, P.S.; Anandham, R.; Han, G.H.; Sa, T. Isolation, characterization, and use for plant growth promotion under salt stress, of ACC deaminase-producing halotolerant bacteria derived from coastal soil. J. Microbiol. Biotechnol. 2010, 20, 1577–1584. [Google Scholar] [CrossRef]
- Beauregard, P.B.; Chai, Y.; Vlamakis, H.; Losick, R.; Kolter, R. Bacillus subtilis biofilm induction by plant polysaccharides. Proc. Natl. Acad. Sci. USA 2013, 110, 1621–1630. [Google Scholar] [CrossRef]
- Shafi, J.; Tian, H.; Ji, M. Bacillus species as versatile weapons for plant pathogens: A review. Biotechnol. Biotechnol. Equip. 2017, 31, 446–459. [Google Scholar] [CrossRef] [Green Version]
- Nephali, L.; Moodley, V.; Piater, L.; Steenkamp, P.; Buthelezi, N.; Dubery, I.; Burgess, K.; Huyser, J.; Tugizimana, F. A Metabolomic Landscape of Maize Plants Treated with a Microbial Biostimulant under Well-Watered and Drought Conditions. Front. Plant Sci. 2021, 12, 676632. [Google Scholar] [CrossRef] [PubMed]
- Nephali, L.; Steenkamp, P.; Burgess, K.; Huyser, J.; Brand, M.; van der Hooft, J.J.J.; Tugizimana, F. Mass Spectral Molecular Networking to Profile the Metabolome of Biostimulant Bacillus Strains. Front. Plant Sci. 2022, 13, 920963. [Google Scholar] [CrossRef] [PubMed]
- Lephatsi, M.; Nephali, L.; Meyer, V.; Piater, L.A.; Buthelezi, N.; Dubery, I.A.; Opperman, H.; Brand, M.; Huyser, J.; Tugizimana, F. Molecular mechanisms associated with microbial biostimulant-mediated growth enhancement, priming and drought stress tolerance in maize plants. Sci. Rep. 2022, 12, 10450. [Google Scholar] [CrossRef]
- Chumthong, A.; Kanjanamaneesathian, M.; Pengnoo, A.; Wiwattanapatapee, R. Water-soluble granules containing Bacillus megaterium for biological control of rice sheath blight: Formulation, bacterial viability and efficacy testing. World J. Microbiol. Biotechnol. 2008, 24, 2499–2507. [Google Scholar] [CrossRef]
- Omer, A.M. Bioformulations of bacillus spores for using as Biofertilizer. Life Sci. J. 2010, 7, 124–131. [Google Scholar]
- Gotor-Vila, A.; Usall, J.; Torres, R.; Abadias, M.; Teixidó, N. Formulation of the biocontrol agent Bacillus amyloliquefaciens CPA-8 using different approaches: Liquid, freeze-drying and fluid-bed spray-drying. BioControl 2017, 62, 545–555. [Google Scholar] [CrossRef]
- Han, L.; Pu, T.; Wang, X.; Liu, B.; Wang, Y.; Feng, J.; Zhang, X. Optimization of a protective medium for enhancing the viability of freeze-dried Bacillus amyloliquefaciens B1408 based on response surface methodology. Cryobiology 2018, 81, 101–106. [Google Scholar] [CrossRef]
- Martínez-Álvarez, J.C.; Castro-Martínez, C.; Sánchez-Peña, P.; Gutiérrez-Dorado, R.; Maldonado-Mendoza, I.E. Development of a powder formulation based on Bacillus cereus sensu lato strain B25 spores for biological control of Fusarium verticillioides in maize plants. World J. Microbiol. Biotechnol. 2016, 32, 75. [Google Scholar] [CrossRef]
- Mahidsanan, T.; Gasaluck, P.; Eumkeb, G. A novel soybean flour as a cryoprotectant in freeze-dried Bacillus subtilis SB-MYP-1. LWT Food Sci. Technol. 2017, 77, 152–159. [Google Scholar] [CrossRef]
- Jayasudha, S.M.; Kirankumar, K.C.; Mesta, R.K.; Ippikoppa, R. Liquid Formulation Using Different Oils and Shelf Life Study of Effective Bacterial Bio-Agents. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 317–324. [Google Scholar] [CrossRef]
- Chung, S.; Lim, J.H.; Kim, S.D. Powder formulation using heat resistant endospores of two multi-functional plant growth promoting rhizobacteria Bacillus strains having phytophtora blight suppression and growth promoting functions. J. Appl. Biol. Chem. 2010, 53, 485–492. [Google Scholar] [CrossRef]
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.R.; Bogdan, A.R.; Miyazawa, M.; Hashimoto, K.; Tsuji, Y. Siderophores in Iron Metabolism: From Mechanism to Therapy Potential. Trends Mol. Med. 2016, 22, 1077–1090. [Google Scholar] [CrossRef]
- Sabir, S.; Arshad, M.; Chaudhari, S.K. Zinc oxide nanoparticles for revolutionizing agriculture: Synthesis and applications. Sci. World J. 2014, 2014, 925494. [Google Scholar] [CrossRef] [PubMed]
- Arkhipova, T.N.; Veselov, S.U.; Melentiev, A.I.; Martynenko, E.V.; Kudoyarova, G.R. Ability of bacterium Bacillus subtilis to produce cytokinins and to influence the growth and endogenous hormone content of lettuce plants. Plant Soil 2005, 272, 201–209. [Google Scholar] [CrossRef]
- Ryu, C.M.; Faragt, M.A.; Hu, C.H.; Reddy, M.S.; Wei, H.X.; Paré, P.W.; Kloepper, J.W. Bacterial volatiles promote growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 4927–4932. [Google Scholar] [CrossRef]
- Zhang, H.; Kim, M.S.; Krishnamachari, V.; Payton, P.; Sun, Y.; Grimson, M.; Farag, M.A.; Ryu, C.M.; Allen, R.; Melo, I.S.; et al. Rhizobacterial volatile emissions regulate auxin homeostasis and cell expansion in Arabidopsis. Planta 2007, 226, 839–851. [Google Scholar] [CrossRef]
- Xie, S.S.; Wu, H.J.; Zang, H.Y.; Wu, L.M.; Zhu, Q.Q.; Gao, X.W. Plant growth promotion by spermidine-producing Bacillus subtilis OKB105. Mol. Plant-Microbe Interact. 2014, 27, 655–663. [Google Scholar] [CrossRef]
- Sasse, J.; Martinoia, E.; Northen, T. Feed Your Friends: Do Plant Exudates Shape the Root Microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, S.S. Impact of Interactions between Rhizosphere and Rhizobacteria: A Review Plant Microbes interaction View project Soil Bioremediation: An approach towards sustainable cleaner technology View project. J. Bacteriol. Mycol. 2018, 5, 1058. [Google Scholar]
- Odoh, C.K. Plant Growth Promoting Rhizobacteria (PGPR): A Bioprotectant bioinoculant for Sustainable Agrobiology. A Review. Int. J. Adv. Res. Biol. Sci. 2017, 4, 123–142. [Google Scholar]
- Poveda, J.; González-Andrés, F. Bacillus as a source of phytohormones for use in agriculture. Appl. Microbiol. Biotechnol. 2021, 105, 8629–8645. [Google Scholar] [CrossRef]
- Shao, J.; Li, S.; Zhang, N.; Cui, X.; Zhou, X.; Zhang, G.; Shen, Q.; Zhang, R. Analysis and cloning of the synthetic pathway of the phytohormone indole-3-acetic acid in the plant-beneficial Bacillus amyloliquefaciens SQR9. Microb. Cell Fact. 2015, 14, 130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shameer, S.; Prasad, T.N.V.K.V. Plant growth promoting rhizobacteria for sustainable agricultural practices with special reference to biotic and abiotic stresses. Plant Growth Regul. 2018, 84, 603–615. [Google Scholar] [CrossRef]
- Wagi, S.; Ahmed, A. Bacillus spp.: Potent microfactories of bacterial IAA. PeerJ 2019, 7, e7258. [Google Scholar] [CrossRef]
- Karadeniz, A.; Topcuoǧlu, Ş.F.; Inan, S. Auxin, gibberellin, cytokinin and abscisic acid production in some bacteria. World J. Microbiol. Biotechnol. 2006, 22, 1061–1064. [Google Scholar] [CrossRef]
- Kudoyarova, G.R.; Korobova, A.V.; Akhiyarova, G.R.; Arkhipova, T.N.; Zaytsev, D.Y.; Prinsen, E.; Egutkin, N.L.; Medvedev, S.S.; Veselov, S.Y. Accumulation of cytokinins in roots and their export to the shoots of durum wheat plants treated with the protonophore carbonyl cyanide m-chlorophenylhydrazone (CCCP). J. Exp. Bot. 2014, 65, 2287–2294. [Google Scholar] [CrossRef] [PubMed]
- Rizza, A.; Jones, A.M. The makings of a gradient: Spatiotemporal distribution of gibberellins in plant development. Curr. Opin. Plant Biol. 2019, 47, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Radhakrishnan, R.; Lee, I.J. Gibberellins producing Bacillus methylotrophicus KE2 supports plant growth and enhances nutritional metabolites and food values of lettuce. Plant Physiol. Biochem. 2016, 109, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, S.; Mulla, S.I.; Lee, K.J.; Chae, J.C.; Shukla, P. VOCs-mediated hormonal signaling and crosstalk with plant growth promoting microbes. Crit. Rev. Biotechnol. 2018, 38, 1277–1296. [Google Scholar] [CrossRef] [PubMed]
- Lemfack, M.C.; Nickel, J.; Dunkel, M.; Preissner, R.; Piechulla, B. MVOC: A database of microbial volatiles. Nucleic Acids Res. 2014, 42, 744–748. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.S.; Dutta, S.; Ann, M.; Raaijmakers, J.M.; Park, K. Promotion of plant growth by Pseudomonas fluorescens strain SS101 via novel volatile organic compounds. Biochem. Biophys. Res. Commun. 2015, 461, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Kanchiswamy, C.N.; Malnoy, M.; Maffei, M.E. Chemical diversity of microbial volatiles and their potential for plant growth and productivity. Front. Plant Sci. 2015, 6, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.Y.; Lu, H.; Wang, X.; Zhang, K.Q.; Li, G.H. Effect of Volatile Organic Compounds from Bacteria on Nematodes. Chem. Biodivers. 2015, 12, 1415–1421. [Google Scholar] [CrossRef]
- Kai, M. Diversity and Distribution of Volatile Secondary Metabolites Throughout Bacillus subtilis Isolates. Front. Microbiol. 2020, 11, 559. [Google Scholar] [CrossRef] [PubMed]
- Piechulla, B.; Lemfack, M.C.; Kai, M. Effects of discrete bioactive microbial volatiles on plants and fungi. Plant Cell Environ. 2017, 40, 2042–2067. [Google Scholar] [CrossRef]
- Rybakova, D.; Rack-Wetzlinger, U.; Cernava, T.; Schaefer, A.; Schmuck, M.; Berg, G. Aerial warfare: A volatile dialogue between the plant pathogen Verticillium longisporum and its antagonist Paenibacillus polymyxa. Front. Plant Sci. 2017, 8, 1294. [Google Scholar] [CrossRef]
- Farag, M.A.; Ryu, C.M.; Sumner, L.W.; Paré, P.W. GC-MS SPME profiling of rhizobacterial volatiles reveals prospective inducers of growth promotion and induced systemic resistance in plants. Phytochemistry 2006, 67, 2262–2268. [Google Scholar] [CrossRef]
- Tahir, H.A.S.; Gu, Q.; Wu, H.; Raza, W.; Hanif, A.; Wu, L.; Colman, M.V.; Gao, X. Plant growth promotion by volatile organic compounds produced by Bacillus subtilis SYST2. Front. Microbiol. 2017, 8, 171. [Google Scholar] [CrossRef]
- Fincheira, P.; Venthur, H.; Mutis, A.; Parada, M.; Quiroz, A. Growth promotion of Lactuca sativa in response to volatile organic compounds emitted from diverse bacterial species. Microbiol. Res. 2016, 193, 39–47. [Google Scholar] [CrossRef]
- Choi, H.K.; Song, G.C.; Yi, H.S.; Ryu, C.M. Field Evaluation of the Bacterial Volatile Derivative 3-Pentanol in Priming for Induced Resistance in Pepper. J. Chem. Ecol. 2014, 40, 882–892. [Google Scholar] [CrossRef] [PubMed]
- Prithiviraj, B.; Perry, L.G.; Badri, D.V.; Vivanco, J.M. Chemical facilitation and induced pathogen resistance mediated by a root-secreted phytotoxin. New Phytol. 2007, 173, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Carvalhais, L.C.; Muzzi, F.; Tan, C.H.; Hsien-Choo, J.; Schenk, P.M. Plant growth in Arabidopsis is assisted by compost soil-derived microbial communities. Front. Plant Sci. 2013, 4, 235. [Google Scholar] [CrossRef] [Green Version]
- Chagas, F.O.; Pessotti, R.D.C.; Caraballo-Rodríguez, A.M.; Pupo, M.T. Chemical signaling involved in plant-microbe interactions. Chem. Soc. Rev. 2018, 47, 1652–1704. [Google Scholar] [CrossRef] [PubMed]
- Bi, S.; Sourjik, V. Stimulus sensing and signal processing in bacterial chemotaxis. Curr. Opin. Microbiol. 2018, 45, 22–29. [Google Scholar] [CrossRef]
- Ankati, S.; Podile, A.R. Metabolites in the root exudates of groundnut change during interaction with plant growth promoting rhizobacteria in a strain-specific manner. J. Plant Physiol. 2019, 243, 153057. [Google Scholar] [CrossRef]
- Feng, H.; Zhang, N.; Fu, R.; Liu, Y.; Krell, T.; Du, W.; Shao, J.; Shen, Q.; Zhang, R. Recognition of dominant attractants by key chemoreceptors mediates recruitment of plant growth-promoting rhizobacteria. Environ. Microbiol. 2018, 21, 402–415. [Google Scholar] [CrossRef]
- Yuan, J.; Zhang, N.; Huang, Q.; Raza, W.; Li, R.; Vivanco, J.M.; Shen, Q. Organic acids from root exudates of banana help root colonization of PGPR strain Bacillus amyloliquefaciens NJN-6. Sci. Rep. 2015, 5, 13438. [Google Scholar] [CrossRef] [PubMed]
- Cesari, A.; Paulucci, N.; López-Gómez, M.; Hidalgo-Castellanos, J.; Plá, C.L.; Dardanelli, M.S. Restrictive water condition modifies the root exudates composition during peanut-PGPR interaction and conditions early events, reversing the negative effects on plant growth. Plant Physiol. Biochem. 2019, 142, 519–527. [Google Scholar] [CrossRef]
- Xie, X.; He, Z.; Chen, N.; Tang, Z.; Wang, Q.; Cai, Y. The Roles of Environmental Factors in Regulation of Oxidative Stress in Plant. Biomed. Res. Int. 2019, 2019, 21–27. [Google Scholar] [CrossRef]
- Hashem, A.; Tabassum, B.; Fathi Abd_Allah, E. Bacillus subtilis: A plant-growth promoting rhizobacterium that also impacts biotic stress. Saudi J. Biol. Sci. 2019, 26, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- Cazorla, F.M.; Romero, D.; Pérez-García, A.; Lugtenberg, B.J.J.; De Vicente, A.; Bloemberg, G. Isolation and characterization of antagonistic Bacillus subtilis strains from the avocado rhizoplane displaying biocontrol activity. J. Appl. Microbiol. 2007, 103, 1950–1959. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Liang, Y.; Wu, M.; Chen, Z.; Lin, J.; Yang, L. Natural products from Bacillus subtilis with antimicrobial properties. Chin. J. Chem. Eng. 2015, 23, 744–754. [Google Scholar] [CrossRef]
- Kinsinger, R.F.; Shirk, M.C.; Fall, R. Rapid surface motility in Bacillus subtilis is dependent on extracellular surfactin and potassium ion. J. Bacteriol. 2003, 185, 5627–5631. [Google Scholar] [CrossRef]
- Yasmin, S.; Zaka, A.; Imran, A.; Zahid, M.A.; Yousaf, S.; Rasul, G.; Arif, M.; Mirza, M.S. Plant growth promotion and suppression of bacterial leaf blight in rice by inoculated bacteria. PLoS ONE 2016, 11, e0160688. [Google Scholar] [CrossRef] [PubMed]
- Zebelo, S.; Song, Y.; Kloepper, J.W.; Fadamiro, H. Rhizobacteria activates (+)-δ-cadinene synthase genes and induces systemic resistance in cotton against beet armyworm (Spodoptera exigua). Plant Cell Environ. 2016, 39, 935–943. [Google Scholar] [CrossRef]
- Pršić, J.; Ongena, M. Elicitors of Plant Immunity Triggered by Beneficial Bacteria. Front. Plant Sci. 2020, 11, 594530. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef]
- Geudens, N.; Martins, J.C. Cyclic lipodepsipeptides from Pseudomonas spp.—Biological Swiss-Army knives. Front. Microbiol. 2018, 9, 1867. [Google Scholar] [CrossRef]
- Chowdhury, S.P.; Uhl, J.; Grosch, R.; Alquéres, S.; Pittroff, S.; Dietel, K.; Schmitt-Kopplin, P.; Borriss, R.; Hartmann, A. Cyclic lipopeptides of Bacillus amyloliquefaciens subsp. plantarum colonizing the lettuce rhizosphere enhance plant defense responses toward the bottom rot pathogen Rhizoctonia solani. Mol. Plant-Microbe Interact. 2015, 28, 984–995. [Google Scholar] [CrossRef]
- Ongena, M.; Jourdan, E.; Adam, A.; Paquot, M.; Brans, A.; Joris, B.; Arpigny, J.L.; Thonart, P. Surfactin and fengycin lipopeptides of Bacillus subtilis as elicitors of induced systemic resistance in plants. Environ. Microbiol. 2007, 9, 1084–1090. [Google Scholar] [CrossRef] [PubMed]
- García-Gutiérrez, L.; Zeriouh, H.; Romero, D.; Cubero, J.; de Vicente, A.; Pérez-García, A. The antagonistic strain Bacillus subtilis UMAF6639 also confers protection to melon plants against cucurbit powdery mildew by activation of jasmonate- and salicylic acid-dependent defence responses. Microb. Biotechnol. 2013, 6, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Cawoy, H.; Mariutto, M.; Henry, G.; Fisher, C.; Vasilyeva, N.; Thonart, P.; Dommes, J.; Ongena, M. Plant defense stimulation by natural isolates of Bacillus depends on efficient surfactin production. Mol. Plant-Microbe Interact. 2014, 27, 87–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, J.; Tonelli, M.L.; Figueredo, M.S.; Ibáñez, F.; Fabra, A. The lipopeptide surfactin triggers induced systemic resistance and priming state responses in Arachis hypogaea L. Eur. J. Plant Pathol. 2018, 152, 845–851. [Google Scholar] [CrossRef]
- Park, K.; Park, Y.S.; Ahamed, J.; Dutta, S.; Ryu, H.; Lee, S.H.; Balaraju, K.; Manir, M.; Moon, S.S. Elicitation of induced systemic resistance of chili pepper by iturin a analogs derived from Bacillus vallismortis EXTN-1. Can. J. Plant Sci. 2016, 96, 564–570. [Google Scholar] [CrossRef]
- Yang, T.; Rao, Z.; Zhang, X.; Xu, M.; Xu, Z.; Yang, S.T. Improved Production of 2,3-Butanediol in Bacillus amyloliquefaciens by Over-Expression of Glyceraldehyde-3-Phosphate Dehydrogenase and 2,3-butanediol Dehydrogenase. PLoS ONE 2013, 8, e76149. [Google Scholar] [CrossRef]
- Rudrappa, T.; Biedrzycki, M.L.; Kunjeti, G.; Donofrio, N.M.; Czymmek, K.J. The rhizobacterial elicitor acetoin induces systemic resistance in Arabidopsis thaliana. Commun. Integr. Biol. 2010, 3, 130–138. [Google Scholar] [CrossRef]
- Kierul, K.; Voigt, B.; Albrecht, D.; Chen, X.H.; Carvalhais, L.C.; Borriss, R. Influence of root exudates on the extracellular proteome of the plant growth-promoting bacterium Bacillus amyloliquefaciens FZB42. Microbiololy 2015, 161, 131–147. [Google Scholar] [CrossRef]
- Tiwari, S.; Prasad, V.; Chauhan, P.S.; Lata, C. Bacillus amyloliquefaciens confers tolerance to various abiotic stresses and modulates plant response to phytohormones through osmoprotection and gene expression regulation in rice. Front. Plant Sci. 2017, 8, 1510. [Google Scholar] [CrossRef]
- Meena, K.K.; Sorty, A.M.; Bitla, U.M.; Choudhary, K.; Gupta, P.; Pareek, A.; Singh, D.P.; Prabha, R.; Sahu, P.K.; Gupta, V.K.; et al. Abiotic stress responses and microbe-mediated mitigation in plants: The omics strategies. Front. Plant Sci. 2017, 8, 172. [Google Scholar] [CrossRef]
- Santoyo, G.; del Orozco-Mosqueda, M.C.; Govindappa, M. Mechanisms of biocontrol and plant growth-promoting activity in soil bacterial species of Bacillus and Pseudomonas: A review. Biocontrol Sci. Technol. 2012, 22, 855–872. [Google Scholar] [CrossRef]
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd-Allah, E.F.; Hashem, A. Phytohormones and beneficial microbes: Essential components for plants to balance stress and fitness. Front. Microbiol. 2017, 8, 2104. [Google Scholar] [CrossRef]
- Sorokan, A.; Veselova, S.; Benkovskaya, G.; Maksimov, I. Endophytic strain Bacillus subtilis 26D increases levels of phytohormones and repairs growth of potato plants after colorado potato beetle damage. Plants 2021, 10, 923. [Google Scholar] [CrossRef]
- Hussain, A.; Hasnain, S. Cytokinin production by some bacteria: Its impact on cell division in cucumber cotyledons. Afr. J. Microbiol. Res. 2009, 3, 704–712. [Google Scholar]
- Arkhipova, T.N.; Prinsen, E.; Veselov, S.U.; Martinenko, E.V.; Melentiev, A.I.; Kudoyarova, G.R. Cytokinin producing bacteria enhance plant growth in drying soil. Plant Soil 2007, 292, 305–315. [Google Scholar] [CrossRef]
- Liu, F.; Xing, S.; Ma, H.; Du, Z.; Ma, B. Cytokinin-producing, plant growth-promoting rhizobacteria that confer resistance to drought stress in Platycladus orientalis container seedlings. Appl. Microbiol. Biotechnol. 2013, 97, 9155–9164. [Google Scholar] [CrossRef]
- Ji, C.; Wang, X.; Tian, H.; Hao, L.; Wang, C.; Zhou, Y.; Xu, R.; Song, X.; Liu, Y.; Du, J.; et al. Effects of Bacillus methylotrophicus M4-1 on physiological and biochemical traits of wheat under salinity stress. J. Appl. Microbiol. 2020, 129, 695–711. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, K.; Zhang, J.; Zhang, Y.; Xu, K.; Yu, D.; Wang, J.; Hu, L.; Chen, L.; Li, C. IAA producing Bacillus altitudinis alleviates iron stress in Triticum aestivum L. seedling by both bioleaching of iron and up-regulation of genes encoding ferritins. Plant Soil 2017, 419, 1–11. [Google Scholar] [CrossRef]
- Fahad, S.; Hussain, S.; Bano, A.; Saud, S.; Hassan, S.; Shan, D.; Khan, F.A.; Khan, F.; Chen, Y.; Wu, C.; et al. Potential role of phytohormones and plant growth-promoting rhizobacteria in abiotic stresses: Consequences for changing environment. Environ. Sci. Pollut. Res. 2015, 22, 4907–4921. [Google Scholar] [CrossRef]
- Pandya, N.D.; Desai, P.V. Gibberellic acid production by Bacillus cereus isolated from the rhizosphere of sugarcane. J. Pure Appl. Microbiol. 2013, 7, 3239–3242. [Google Scholar]
- Kang, S.M.; Khan, A.L.; Waqas, M.; Asaf, S.; Lee, K.E.; Park, Y.G.; Kim, A.Y.; Khan, M.A.; You, Y.H.; Lee, I.J. Integrated phytohormone production by the plant growth-promoting rhizobacterium Bacillus tequilensis SSB07 induced thermotolerance in soybean. J. Plant Interact. 2019, 14, 416–423. [Google Scholar] [CrossRef]
- Wilkinson, S.; Kudoyarova, G.R.; Veselov, D.S.; Arkhipova, T.N.; Davies, W.J. Plant hormone interactions: Innovative targets for crop breeding and management. J. Exp. Bot. 2012, 63, 3499–3509. [Google Scholar] [CrossRef]
- Chaves, M.M.; Maroco, J.P.; Pereira, J.S. Understanding plant responses to drought—From genes to the whole plant. Funct. Plant Biol. 2003, 30, 239–264. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Jia, W.; Yang, J.; Ismail, A.M. Role of ABA in integrating plant responses to drought and salt stresses. Field Crop. Res. 2006, 97, 111–119. [Google Scholar] [CrossRef]
- Pan, W.; Lu, Q.; Xu, Q.R.; Zhang, R.R.; Li, H.Y.; Yang, Y.H.; Liu, H.J.; Du, S.T. Abscisic acid-generating bacteria can reduce Cd concentration in pakchoi grown in Cd-contaminated soil. Ecotoxicol. Environ. Saf. 2019, 177, 100–107. [Google Scholar] [CrossRef]
- Zhang, H.; Xie, X.; Kim, M.S.; Kornyeyev, D.A.; Holaday, S.; Paré, P.W. Soil bacteria augment Arabidopsis photosynthesis by decreasing glucose sensing and abscisic acid levels in planta. Plant J. 2008, 56, 264–273. [Google Scholar] [CrossRef]
- Zhang, H.; Murzello, C.; Sun, Y.; Kim, M.S.; Xie, X.; Jeter, R.M.; Zak, J.C.; Dowd, S.E.; Paré, P.W. Choline and osmotic-stress tolerance induced in arabidopsis by the soil microbe Bacillus subtilis (GB03). Mol. Plant-Microbe Interact. 2010, 23, 1097–1104. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, D.; Hanson, A.D. Compounds in Higher Plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1993, 44, 357–384. [Google Scholar] [CrossRef]
- Nephali, L.; Piater, L.A.; Dubery, I.A.; Patterson, V.; Huyser, J.; Burgess, K.; Tugizimana, F. Biostimulants for plant growth and mitigation of abiotic stresses: A metabolomics perspective. Metabolites 2020, 10, 505. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.G.; Mun, B.G.; Kang, S.M.; Hussain, A.; Shahzad, R.; Seo, C.W.; Kim, A.Y.; Lee, S.U.; Oh, K.Y.; Lee, D.Y.; et al. Bacillus aryabhattai SRB02 tolerates oxidative and nitrosative stress and promotes the growth of soybean by modulating the production of phytohormones. PLoS ONE 2017, 12, e0173203. [Google Scholar] [CrossRef] [PubMed]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef]
- Othibeng, K.; Nephali, L.; Myoli, A.; Buthelezi, N.; Jonker, W.; Huyser, J.; Tugizimana, F. Metabolic Circuits in Sap Extracts Reflect the Effects of a Microbial Biostimulant on Maize Metabolism under Drought Conditions. Plants 2022, 11, 510. [Google Scholar] [CrossRef] [PubMed]
- Hasin, Y.; Seldin, M.; Lusis, A. Multi-omics approaches to disease. Genome Biol. 2017, 18, 83. [Google Scholar] [CrossRef]
- Baek, D.; Rokibuzzaman, M.; Khan, A.; Kim, M.C.; Park, H.J.; Yun, D.J.; Chung, Y.R. Plant-Growth Promoting Bacillus oryzicola YC7007 Modulates Stress-Response Gene Expression and Provides Protection from Salt Stress. Front. Plant Sci. 2020, 10, 1646. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, P.S.; Lata, C.; Tiwari, S.; Chauhan, A.S.; Mishra, S.K.; Agrawal, L.; Chakrabarty, D.; Nautiyal, C.S. Transcriptional alterations reveal Bacillus amyloliquefaciens-rice cooperation under salt stress. Sci. Rep. 2019, 9, 11912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, S.; Souleimanov, A.; Smith, D.L. Thuricin17 Production and Proteome Differences in Bacillus thuringiensis NEB17 Cell-Free Supernatant under NaCl Stress. Front. Sustain. Food Syst. 2021, 5, 630628. [Google Scholar] [CrossRef]
- Hall, R.D.; D’Auria, J.C.; Silva Ferreira, A.C.; Gibon, Y.; Kruszka, D.; Mishra, P.; van de Zedde, R. High-throughput plant phenotyping: A role for metabolomics? Trends Plant Sci. 2022, 27, 549–563. [Google Scholar] [CrossRef]
- van Dam, N.M.; Bouwmeester, H.J. Metabolomics in the Rhizosphere: Tapping into Belowground Chemical Communication. Trends Plant Sci. 2016, 21, 256–265. [Google Scholar] [CrossRef]
- Comolli, L.R. Intra- and inter-species interactions in microbial communities. Front. Microbiol. 2014, 5, 629. [Google Scholar] [CrossRef]
- Pascale, A.; Proietti, S.; Pantelides, I.S.; Stringlis, I.A. Modulation of the Root Microbiome by Plant Molecules: The Basis for Targeted Disease Suppression and Plant Growth Promotion. Front. Plant Sci. 2020, 10, 1741. [Google Scholar] [CrossRef]
- Vishwakarma, K.; Kumar, N.; Shandilya, C.; Mohapatra, S.; Bhayana, S.; Varma, A. Revisiting Plant–Microbe Interactions and Microbial Consortia Application for Enhancing Sustainable Agriculture: A Review. Front. Microbiol. 2020, 11, 560406. [Google Scholar] [CrossRef] [PubMed]
- Yin, C.; Casa Vargas, J.M.; Schlatter, D.C.; Hagerty, C.H.; Hulbert, S.H.; Paulitz, T.C. Rhizosphere community selection reveals bacteria associated with reduced root disease. Microbiome 2021, 9, 86. [Google Scholar] [CrossRef] [PubMed]
- Nyholm, L.; Koziol, A.; Marcos, S.; Botnen, A.B.; Aizpurua, O.; Gopalakrishnan, S.; Limborg, M.T.; Gilbert, M.T.P.; Alberdi, A. Holo-Omics: Integrated Host-Microbiota Multi-omics for Basic and Applied Biological Research. iScience 2020, 23, 101414. [Google Scholar] [CrossRef]
- Li, D.; Carr, G.; Zhang, Y.; Williams, D.E.; Amlani, A.; Bottriell, H.; Mui, A.L.F.; Andersen, R.J. Turnagainolides A and B, cyclic depsipeptides produced in culture by a Bacillus sp.: Isolation, structure elucidation, and synthesis. J. Nat. Prod. 2011, 74, 1093–1099. [Google Scholar] [CrossRef]
- Tareq, F.S.; Shin, H.J. Bacilotetrins A and B, Anti-Staphylococcal Cyclic-Lipotetrapeptides from a Marine-Derived Bacillus subtilis. J. Nat. Prod. 2017, 80, 2889–2892. [Google Scholar] [CrossRef]
- Ravu, R.R.; Jacob, M.R.; Chen, X.; Wang, M.; Nasrin, S.; Kloepper, J.W.; Liles, M.R.; Mead, D.A.; Khan, I.A.; Li, X.C. Bacillusin A, an antibacterial macrodiolide from Bacillus amyloliquefaciens AP183. J. Nat. Prod. 2015, 78, 924–928. [Google Scholar] [CrossRef] [PubMed]
- Tareq, F.S.; Lee, M.A.; Lee, H.S.; Lee, J.S.; Lee, Y.J.; Shin, H.J. Gageostatins A-C, antimicrobial linear lipopeptides from a marine Bacillus subtilis. Mar. Drugs 2014, 12, 871–885. [Google Scholar] [CrossRef] [PubMed]
- Romero-Tabarez, M.; Jansen, R.; Sylla, M.; Lünsdorf, H.; Häußler, S.; Santosa, D.A.; Timmis, K.N.; Molinari, G. 7-O-malonyl macrolactin A, a new macrolactin antibiotic from Bacillus subtilis active against methicillin-resistant Staphylococcus aureus, Vancomycin-resistant enterococci, and a small-colony variant of Burkholderia cepacia. Antimicrob. Agents Chemother. 2006, 50, 1701–1709. [Google Scholar] [CrossRef]
- Ma, Z.; Hu, J. Plipastatin A1 produced by a marine sediment-derived Bacillus amyloliquefaciens SH-B74 contributes to the control of gray mold disease in tomato. 3 Biotech 2018, 8, 125. [Google Scholar] [CrossRef]
- Chakraborty, K.; Thilakan, B.; Raola, V.K. Antimicrobial polyketide furanoterpenoids from seaweed-associated heterotrophic bacterium Bacillus subtilis MTCC 10403. Phytochemistry 2017, 142, 112–125. [Google Scholar] [CrossRef]
- Xie, C.L.; Xia, J.M.; Su, R.Q.; Li, J.; Liu, Y.; Yang, X.W.; Yang, Q. Bacilsubteramide A, a new indole alkaloid, from the deep-sea-derived Bacillus subterraneus 11593. Nat. Prod. Res. 2018, 32, 2553–2557. [Google Scholar] [CrossRef] [PubMed]
- Pinzón-Espinosa, A.; Martinez-Matamoros, D.; Castellanos, L.; Duque, C.; Rodríguez, J.; Jiménez, C.; Ramos, F.A. Cereusitin A, a cyclic tetrapeptide from a Bacillus cereus strain isolated from the soft coral Antillogorgia (syn. Pseudopterogorgia) elisabethae. Tetrahedron Lett. 2017, 58, 634–637. [Google Scholar] [CrossRef]
- Jin, J.; Lao, J.; Zhou, R.; He, W.; Qin, Y.; Zhong, C.; Xie, J.; Liu, H.; Wan, D.; Zhang, S.; et al. Simultaneous identification and dynamic analysis of saccharides during steam processing of rhizomes of Polygonatum cyrtonema by HPLC–QTOF–MS/MS. Molecules 2018, 23, 2855. [Google Scholar] [CrossRef]
- Anjum, K.; Bi, H.; Chai, W.; Lian, X.Y.; Zhang, Z. Antiglioma pseurotin A from marine Bacillus sp. FS8D regulating tumour metabolic enzymes. Nat. Prod. Res. 2018, 32, 1353–1356. [Google Scholar] [CrossRef] [PubMed]
- Le Marrec, C.; Hyronimus, B.; Bressollier, P.; Verneuil, B.; Urdaci, M.C. Biochemical and genetic characterization of coagulin, a new antilisterial bacteriocin in the pediocin family of bacteriocins, produced by Bacillus coagulans I4. Appl. Environ. Microbiol. 2000, 66, 5213–5220. [Google Scholar] [CrossRef] [Green Version]
- Kamoun, F.; Mejdoub, H.; Aouissaoui, H.; Reinbolt, J.; Hammami, A.; Jaoua, S. Purification, amino acid sequence and characterization of Bacthuricin F4, a new bacteriocin produced by Bacillus thuringiensis. J. Appl. Microbiol. 2005, 98, 881–888. [Google Scholar] [CrossRef]
- Bizani, D.; Dominguez, A.P.M.; Brandelli, A. Purification and partial chemical characterization of the antimicrobial peptide cerein 8A. Lett. Appl. Microbiol. 2005, 41, 269–273. [Google Scholar] [CrossRef]
- Lisboa, M.P.; Bonatto, D.; Bizani, D.; Henriques, J.A.P.; Brandelli, A. Characterization of a bacteriocin-like substance produced by Bacillus amyloliquefaciens isolated from the Brazilian Atlantic forest. Int. Microbiol. 2006, 9, 111–118. [Google Scholar]
- Chehimi, S.; Delalande, F.; Sablé, S.; Hajlaoui, M.R.; Van Dorsselaer, A.; Limam, F.; Pons, A.M. Purification and partial amino acid sequence of thuricin S, a new anti-Listeria bacteriocin from Bacillus thuringiensis. Can. J. Microbiol. 2007, 53, 284–290. [Google Scholar] [CrossRef]
- Kalinovskaya, N.I.; Kuznetsova, T.A.; Ivanova, E.P.; Voinov, V.G.; Huth, F.; Laatsch, H. Characterization of Surfactin-like Cyclic Depsipeptides Synthesized by Bacillus pumilus from Ascidian Halocynthia aurantium. Mar. Biotechnol. 2002, 4, 179–188. [Google Scholar] [CrossRef]
- Steinborn, G.; Hajirezaei, M.R.; Hofemeister, J. bac genes for recombinant bacilysin and anticapsin production in Bacillus host strains. Arch. Microbiol. 2005, 183, 71–79. [Google Scholar] [CrossRef]
- Tamehiro, N.; Okamoto-Hosoya, Y.; Okamoto, S.; Ubukata, M.; Hamada, M.; Naganawa, H.; Ochi, K. Bacilysocin, a novel phospholipid antibiotic produced by Bacillus subtilis 168. Antimicrob. Agents Chemother. 2002, 46, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Luzzatto-Knaan, T.; Melnik, A.V.; Dorrestein, P.C. Mass Spectrometry Uncovers the Role of Surfactin as an Interspecies Recruitment Factor. ACS Chem. Biol. 2019, 14, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Watrous, J.; Roach, P.; Alexandrov, T.; Heath, B.S.; Yang, J.Y.; Kersten, R.D.; Van Der Voort, M.; Pogliano, K.; Gross, H.; Raaijmakers, J.M.; et al. Mass spectral molecular networking of living microbial colonies. Proc. Natl. Acad. Sci. USA 2012, 109, 1743–1752. [Google Scholar] [CrossRef] [PubMed]
- Tugizimana, F.; Engel, J.; Salek, R.; Dubery, I.; Piater, L.; Burgess, K. The disruptive 4IR in life sciences: Metanolomics. In The Disruptive Fourth Industrial Revolution; Doorsamy, W., Paul, B.S., Marwala, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2020; pp. 227–256. [Google Scholar]
- Dodds, J.N.; Baker, E.S. Ion Mobility Spectrometry: Fundamental Concepts, Instrumentation, Applications, and the Road Ahead. J. Am. Soc. Mass Spectrom. 2019, 30, 2185–2195. [Google Scholar] [CrossRef] [PubMed]
- Armenta, S.; Esteve-Turrillas, F.A.; Alcalà, M. Analysis of hazardous chemicals by “stand alone” drift tube ion mobility spectrometry: A review. Anal. Methods 2020, 12, 1163–1181. [Google Scholar] [CrossRef]
- Ratiu, I.A.; Bocos-Bintintan, V.; Patrut, A.; Moll, V.H.; Turner, M.; Thomas, C.L.P. Discrimination of bacteria by rapid sensing their metabolic volatiles using an aspiration-type ion mobility spectrometer (a-IMS) and gas chromatography-mass spectrometry GC-MS. Anal. Chim. Acta 2017, 982, 209–217. [Google Scholar] [CrossRef]
- Quinn, R.A.; Nothias, L.F.; Vining, O.; Meehan, M.; Esquenazi, E.; Dorrestein, P.C. Molecular Networking As a Drug Discovery, Drug Metabolism, and Precision Medicine Strategy. Trends Pharmacol. Sci. 2017, 38, 143–154. [Google Scholar] [CrossRef]
- Rogers, S.; Ong, C.W.; Wandy, J.; Ernst, M.; Ridder, L.; Van Der Hooft, J.J.J. Deciphering complex metabolite mixtures by unsupervised and supervised substructure discovery and semi-automated annotation from MS/MS spectra. Faraday Discuss. 2019, 218, 284–302. [Google Scholar] [CrossRef]
- Nothias, L.F.; Petras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-based molecular networking in the GNPS analysis environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef]
- Wang, T.; Lu, Q.; Sun, C.; Lukianov, D.; Osterman, I.A.; Sergiev, P.V.; Dontsova, O.A.; Hu, X.; You, X.; Liu, S.; et al. Hetiamacin E and F, New amicoumacin antibiotics from Bacillus subtilis PJS using MS/MS-based molecular networking. Molecules 2020, 25, 4446. [Google Scholar] [CrossRef] [PubMed]
- Purves, K.; Macintyre, L.; Brennan, D.; Hreggviðsson, G.; Kuttner, E.; Ásgeirsdóttir, M.E.; Young, L.C.; Green, D.H.; Edrada-Ebel, R.; Duncan, K.R. Using molecular networking for microbial secondary metabolite bioprospecting. Metabolites 2016, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Meyer, H.; Weidmann, H.; Lalk, M. Methodological approaches to help unravel the intracellular metabolome of Bacillus subtilis. Microb. Cell Fact. 2013, 12, 69. [Google Scholar] [CrossRef] [PubMed]
- Mondol, M.A.M.; Shin, H.J.; Islam, M.T. Diversity of secondary metabolites from marine bacillus species: Chemistry and biological activity. Mar. Drugs 2013, 11, 2846–2872. [Google Scholar] [CrossRef] [PubMed]
- Shahid, I.; Han, J.; Hanooq, S.; Malik, K.A.; Borchers, C.H.; Mehnaz, S. Profiling of Metabolites of Bacillus spp. and Their Application in Sustainable Plant Growth Promotion and Biocontrol. Front. Sustain. Food Syst. 2021, 5, 605195. [Google Scholar] [CrossRef]
- Nigris, S.; Baldan, E.; Tondello, A.; Zanella, F.; Vitulo, N.; Favaro, G.; Guidolin, V.; Bordin, N.; Telatin, A.; Barizza, E.; et al. Biocontrol traits of Bacillus licheniformis GL174, a culturable endophyte of Vitis vinifera cv. Glera 06 Biological Sciences 0604 Genetics 06 Biological Sciences 0607 Plant Biology 06 Biological Sciences 0605 Microbiology. BMC Microbiol. 2018, 18, 133. [Google Scholar]
- Miljaković, D.; Marinković, J.; Balešević-Tubić, S. The significance of Bacillus spp. in disease suppression and growth promotion of field and vegetable crops. Microorganisms 2020, 8, 1037. [Google Scholar] [CrossRef]
- Tran, C.; Cock, I.E.; Chen, X.; Feng, Y. Antimicrobial Bacillus: Metabolites and Their Mode of Action. Antibiotics 2022, 11, 88. [Google Scholar] [CrossRef]
- Mhlongo, M.I.; Piater, L.A.; Madala, N.E.; Labuschagne, N.; Dubery, I.A.; Hall, R.D. The chemistry of plant–microbe interactions in the rhizosphere and the potential for metabolomics to reveal signaling related to defense priming and induced systemic resistance. Front. Plant Sci. 2018, 9, 112. [Google Scholar] [CrossRef]
- Andrić, S.; Meyer, T.; Ongena, M. Bacillus Responses to Plant-Associated Fungal and Bacterial Communities. Front. Microbiol. 2020, 11, 1350. [Google Scholar] [CrossRef]
- Venturi, V.; Bez, C. A call to arms for cell–cell interactions between bacteria in the plant microbiome. Trends Plant Sci. 2021, 26, 1126–1132. [Google Scholar] [CrossRef] [PubMed]
- Korenblum, E.; Dong, Y.; Szymanski, J.; Panda, S.; Jozwiak, A.; Massalha, H.; Meir, S.; Rogachev, I.; Aharoni, A. Rhizosphere microbiome mediates systemic root metabolite exudation by root-to-root signaling. Proc. Natl. Acad. Sci. USA 2020, 117, 3874–3883. [Google Scholar] [CrossRef] [PubMed]
- Wen, T.; Zhao, M.; Liu, T.; Huang, Q.; Yuan, J.; Shen, Q. High abundance of Ralstonia solanacearum changed tomato rhizosphere microbiome and metabolome. BMC Plant Biol. 2020, 20, 166. [Google Scholar] [CrossRef]
- Andrić, S.; Meyer, T.; Rigolet, A.; Prigent-Combaret, C.; Höfte, M.; Balleux, G.; Steels, S.; Hoff, G.; De Mot, R.; McCann, A.; et al. Lipopeptide Interplay Mediates Molecular Interactions between Soil Bacilli and Pseudomonads. Microbiol. Spectr. 2021, 9, e0203821. [Google Scholar] [CrossRef]
- Kang, W.S.; Chen, L.J.L.J.; Wang, Y.Y.; Zhu, X.F.; Liu, X.Y.; Fan, H.; Duan, Y.X. Bacillus simplex treatment promotes soybean defence against soybean cyst nematodes: A metabolomics study using GC-MS. PLoS ONE 2020, 15, e0237194. [Google Scholar] [CrossRef] [PubMed]
- Carlson, R.; Tugizimana, F.; Steenkamp, P.A.; Dubery, I.A.; Hassen, A.I.; Labuschagne, N. Rhizobacteria-induced systemic resilience in Sorghum bicolor (L.) moench against Fusarium pseudograminearum crown rot under drought stress conditions. Biol. Control 2020, 151, 104395. [Google Scholar] [CrossRef]
Figure 1.
Intra-and extracellular biosynthesis of metal-nanoparticles by Bacillus spp. Extracellular biosynthesis of metal-nanoparticles is carried out by trapping metal ions on the cell surface and reducing them in the presence of secreted enzymes or metabolite and/or membrane-bound enzymes. In the intracellular biosynthesis of metal-nanoparticles, after transfer of metal ions into cell cytoplasm, the metal ions are reduced as a result of metabolic reactions with enzymes such as alpha-NADPH-dependent nitrate reductase.
Figure 1.
Intra-and extracellular biosynthesis of metal-nanoparticles by Bacillus spp. Extracellular biosynthesis of metal-nanoparticles is carried out by trapping metal ions on the cell surface and reducing them in the presence of secreted enzymes or metabolite and/or membrane-bound enzymes. In the intracellular biosynthesis of metal-nanoparticles, after transfer of metal ions into cell cytoplasm, the metal ions are reduced as a result of metabolic reactions with enzymes such as alpha-NADPH-dependent nitrate reductase.

Figure 2.
Some molecules involved in Bacillus-plant interactions. Plants produce various compounds including the hormones salicylic acid (SA), jasmonic acid (JA), cytokinins (CK), and indole acetic acid. Plants also produce organic acids, flavonoids, amino acids, and 1-aminocyclopropane-1-carboxylate (ACC) as signaling molecules. On the other hand, Bacillus produce volatiles such as 2,3 butanediol, lipopeptides such as surfactin, and phytohormones such as gibberellic acid and acyl homoserine lactone (AHL) as signaling molecules.
Figure 2.
Some molecules involved in Bacillus-plant interactions. Plants produce various compounds including the hormones salicylic acid (SA), jasmonic acid (JA), cytokinins (CK), and indole acetic acid. Plants also produce organic acids, flavonoids, amino acids, and 1-aminocyclopropane-1-carboxylate (ACC) as signaling molecules. On the other hand, Bacillus produce volatiles such as 2,3 butanediol, lipopeptides such as surfactin, and phytohormones such as gibberellic acid and acyl homoserine lactone (AHL) as signaling molecules.

Figure 3.
An overview of mechanisms employed by Bacillus spp. in the mitigation of biotic and abiotic stresses. Bacillus produce cyclic lipopetides which activate pathways regulated by jasmonic acid and ethylene and thus elicit induced systemic resistance (ISR). They also produce hydrogen cyanide (HCN) and the enzyme 1-aminocyclopropane-1-carboxylate (ACC) deaminase which lower the levels of plant ethylene. Siderophores produced by Bacillus chelate iron thus making it unavailable to the pathogen. Several root associated Bacillus spp. bacteria produce zeatin, gibberellic acid (GA), indole-3-acetic acid (IAA), salicylic acid (SA), abscisic acid (ABA) as well as volatile organic acids (VOCs) which help plants to withstand stress by enhancing its antioxidant potential, by up-regulation of the antioxidant system and by accumulation of compatible osmolytes thus reducing oxidative stress-induced damage; improving photosynthetic capacity and membrane stability; promoting cell division and stomatal regulation; stimulating growth of root system, and acquisition of water and nutrients.
Figure 3.
An overview of mechanisms employed by Bacillus spp. in the mitigation of biotic and abiotic stresses. Bacillus produce cyclic lipopetides which activate pathways regulated by jasmonic acid and ethylene and thus elicit induced systemic resistance (ISR). They also produce hydrogen cyanide (HCN) and the enzyme 1-aminocyclopropane-1-carboxylate (ACC) deaminase which lower the levels of plant ethylene. Siderophores produced by Bacillus chelate iron thus making it unavailable to the pathogen. Several root associated Bacillus spp. bacteria produce zeatin, gibberellic acid (GA), indole-3-acetic acid (IAA), salicylic acid (SA), abscisic acid (ABA) as well as volatile organic acids (VOCs) which help plants to withstand stress by enhancing its antioxidant potential, by up-regulation of the antioxidant system and by accumulation of compatible osmolytes thus reducing oxidative stress-induced damage; improving photosynthetic capacity and membrane stability; promoting cell division and stomatal regulation; stimulating growth of root system, and acquisition of water and nutrients.

Figure 4.
Omics technologies in systems biology. The omics cascade shows the flow of information from gene level to the metabolome with metabolomics being the closest link to the phenotype.
Figure 4.
Omics technologies in systems biology. The omics cascade shows the flow of information from gene level to the metabolome with metabolomics being the closest link to the phenotype.


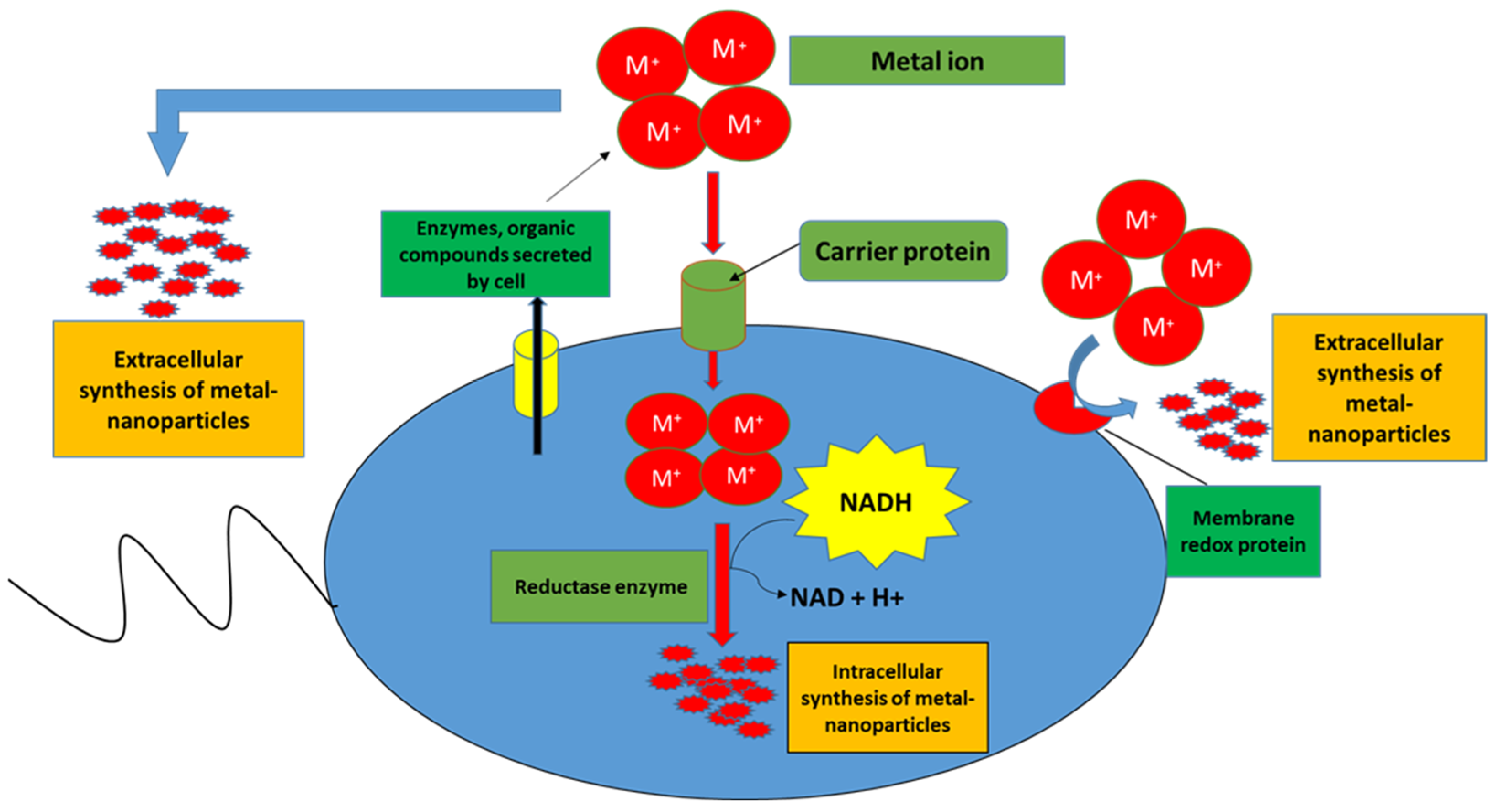{kind=link}
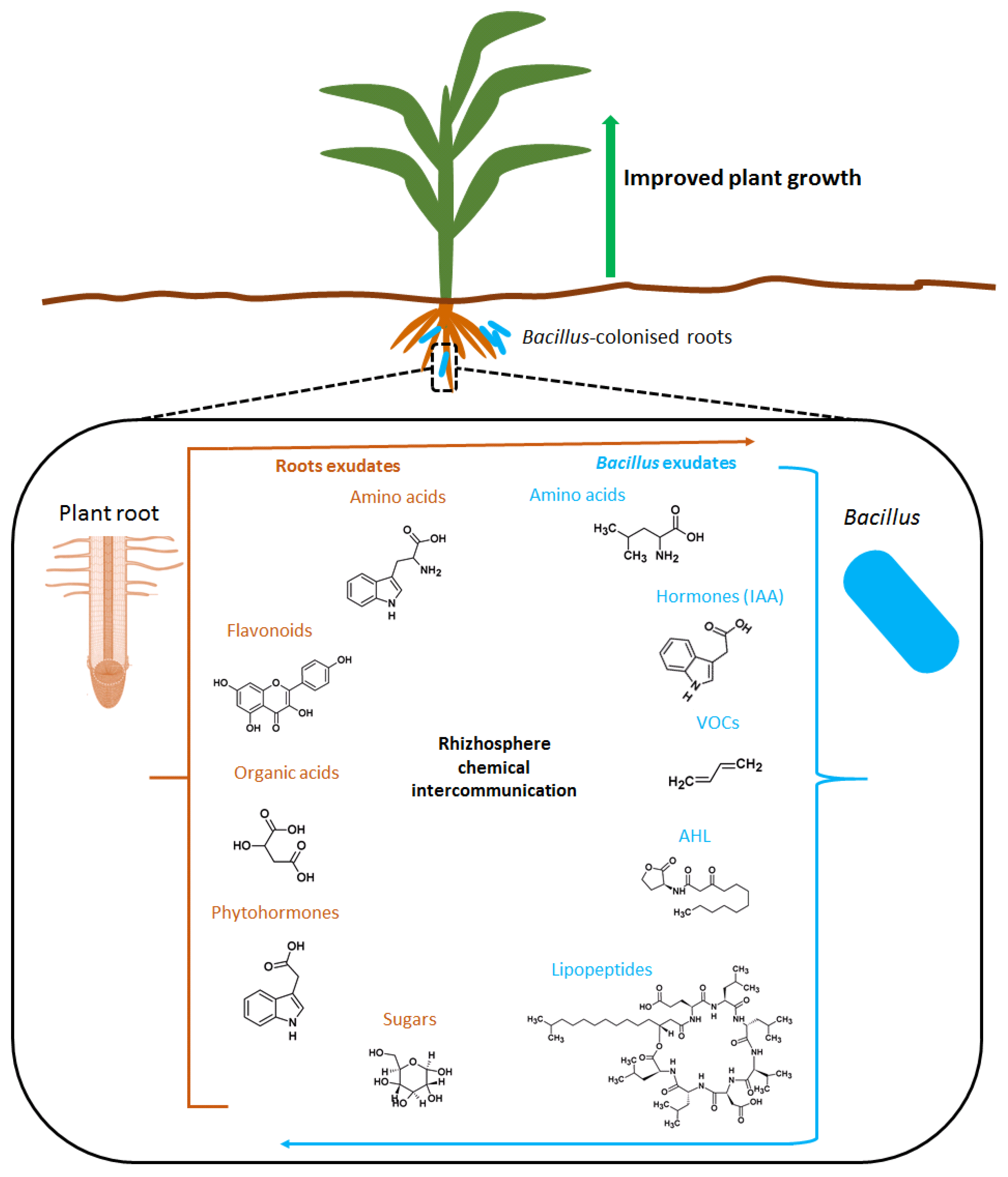{kind=link}
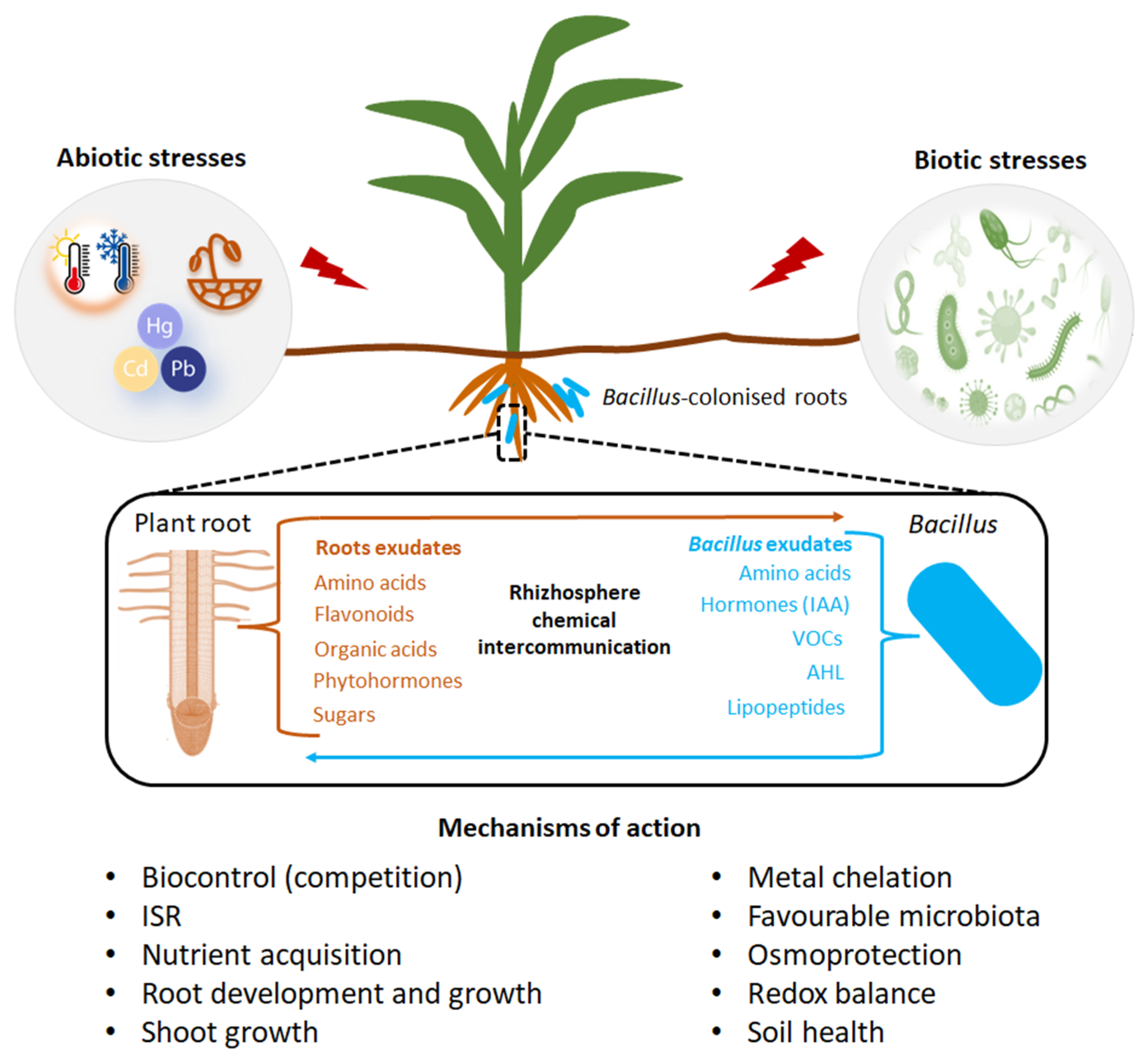{kind=link}
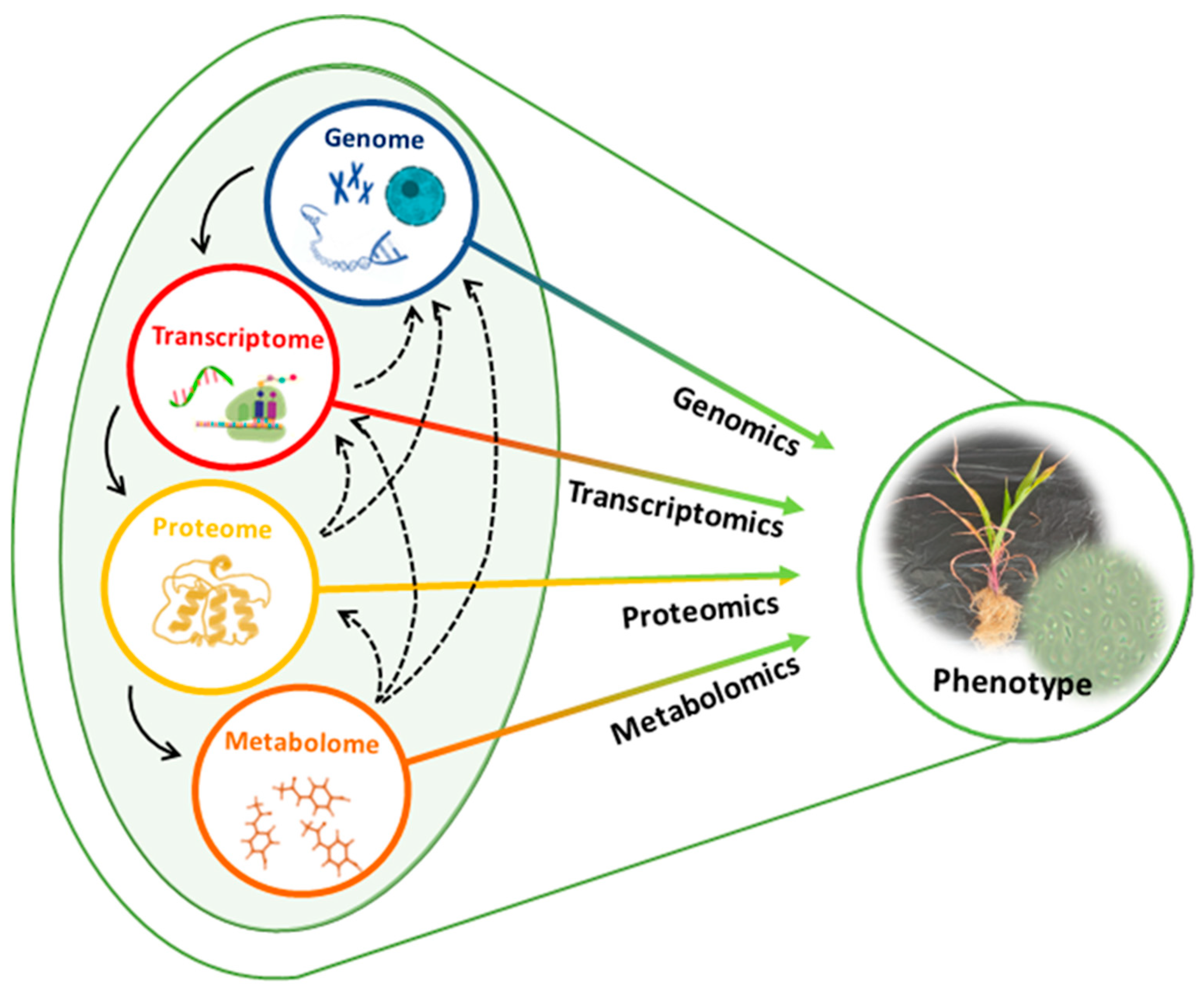{kind=link}
Table 1.
Bacillus spp. biostimulant formulations.
| Microorganism | Formulation Type | Additives | References |
|---|---|---|---|
| Consortium * of Bacillus licheniformis, Bacillus laterosporus and Bacillus amyloliquefaciens | Liquid | No additives disclosed | [11,12,13] |
| Bacillus megaterium | Granules | Lactose monohydrate PVPK-30, sodium alignate | [14] |
| Bacillus megaterium | Powder | Talc, clay and cellulose; CMC, sodium benzoate, CaCO3, Glucose, sucrose, mannitol, yeast, peptone. | [15] |
| Bacillus amyloliquefacines | Powder | Sucrose, powder skimmed milk, MgSO4 | [16] |
| Bacillus amyloliquefacines | Liquid | Sucrose, powder skimmed milk, MgSO4 | [16] |
| Bacillus amyloliquefacines | Powder | MgSO4 | [17] |
| Bacillus cereus | Powder | Glucose, fructose, D-galactose, sucrose, trehalose, cellobiose, glutamic acid, soluble starch, glycerol, sorbitol, peptone, nonfat skimmed milk. | [18] |
| Bacillus cereus | Powder | Talc, CMC, CaCO3, Glucose | [19] |
| Bacillus subtilis | Powder | Soybean flour | [20] |
| Bacillus subtilis | Liquid | Groundnut oil, Pongamia oil and sunflower oil; glycerol | [21] |
| Bacillus subtilis and licheniformis | Powder | Natural zeolite Synthetic zeolite | [21] |
* This consortium is on the market as a biostimulant product, BACSTIM® 100.
Table 2.
Effects of VOCs produced by Bacillus.
| VOCs | Bacillus spp. | Plant | Response | References |
|---|---|---|---|---|
| 2,3-Butandiol | B. subtilis; B. amyloliquenfaciens | Arabidopsis thaliana, Solanum tuberusom | Induced systemic resistance/tolerance, plant growth promotion | [47,48] |
| Butyrolactone | B. cereus, B. subtilis | Arabidopsis thaliana | Growth promotion, modify root system architecture | [49] |
| Acetophenone | B. megaterium, B. pumilis | Arabidopsis thaliana | Growth promotion, modify root system architecture | [49] |
| 2-Butanone | B. subtilis; B. amyloliquenfaciens | Nicotinia tabacum | Growth promotion | [50] |
| 3-Pentanol | B. megaterium, B. pumilis | Capsicum annum | Induced systemic resistance, plant growth promotion | [51] |
Table 3.
Identified metabolites in the intracellular and extracellular chemical space of Bacillus spp.
Table 3.
Identified metabolites in the intracellular and extracellular chemical space of Bacillus spp.
| Bacillus Species | Metabolite | Chemical Nature | Bioactivity | References |
|---|---|---|---|---|
| B. subtilis (unidentified marin starin) | Bacilotetrins | Cyclic-lipotetrapeptides | Antimicrobial | [115] |
| B. amyloliquefaciens AP183 | Bacillusin | Macrocyclic polyene | Antimicrobial | [116] |
| B. subtilis (unidentified marin starin) | Gageotetrins | Linear lipopeptides | Antimicrobial, anticancer | [117] |
| B. subtilis DSM 16696 | Macrolactin | Macrolides | Antimicrobial | [118] |
| B. subtilis DSM 16697 | Plipastatin A | Lipopeptides | Antifungal | [119] |
| B. subtilis MTCC 10403 | Furanoterpenoids | Polyketide | Antimicrobial | [120] |
| Unidentified Bacillus strain | Turnagainolides | Depsipeptides | Activation of SHIP1 | [114] |
| B. subterraneus 11593 | Bacilsubteramide A | Alkaloid | [121] | |
| B. cereus RKHC-09 | Cereusitin A | Cyclic tetrapeptide | Antifungal | [122] |
| B. amyloliquefaciens HAB-2 | Bacillomycin | Cyclic lipopeptide | Antifungal | [123] |
| Bacillus sp. FS8D | Pseurotin A | Spirocyclic | Anticancer | [124] |
| B. coagulans 14 | Coagulin | Peptide | Antibacterial | [125] |
| B. thuringenesis | Bacthurucin f4 | Peptide | Antifungal | [126] |
| B. cereus | Cerein | Peptide | Antibacterial | [127] |
| B. megaterium | Megacin | Peptide | Antibacterial | [128] |
| B. thuringenesis S | Thuricin | Peptide | [129] | |
| B. licheniformis | Halobacillin 5b | Hemolytic, cytotoxic | [130] | |
| B. amyloliquefaciens GSB272 | Bacilysin 1 | Antifungal, antibacterial | [131] | |
| B. subtilis 168 | Bacilysocin | Fungicidal, antibacterial | [132] | |
| B. licheniformis 1001, B. licheniformis M017, B. amyloliquefaciens | Lichenysins Surfactins | antimicrobials | [12] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tsotetsi, T.; Nephali, L.; Malebe, M.; Tugizimana, F. Bacillus for Plant Growth Promotion and Stress Resilience: What Have We Learned? Plants 2022, 11, 2482. https://doi.org/10.3390/plants11192482
AMA Style
Tsotetsi T, Nephali L, Malebe M, Tugizimana F. Bacillus for Plant Growth Promotion and Stress Resilience: What Have We Learned? Plants. 2022; 11(19):2482. https://doi.org/10.3390/plants11192482
Chicago/Turabian StyleTsotetsi, Teboho, Lerato Nephali, Motumiseng Malebe, and Fidele Tugizimana. 2022. "Bacillus for Plant Growth Promotion and Stress Resilience: What Have We Learned?" Plants 11, no. 19: 2482. https://doi.org/10.3390/plants11192482
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.