
Application of Biostimulants in Tomato Plants (Solanum lycopersicum) to Enhance Plant Growth and Salt Stress Tolerance
 ,
,  and
and
Abstract
:1. Introduction
2. Results
2.1. Phenotypic Observations of Biostimulant-Treated Plants under Control and Salt Stress Conditions
2.2. Effects of Biostimulants on Growth Parameters
2.3. Physiological Responses of Biostimulants Application
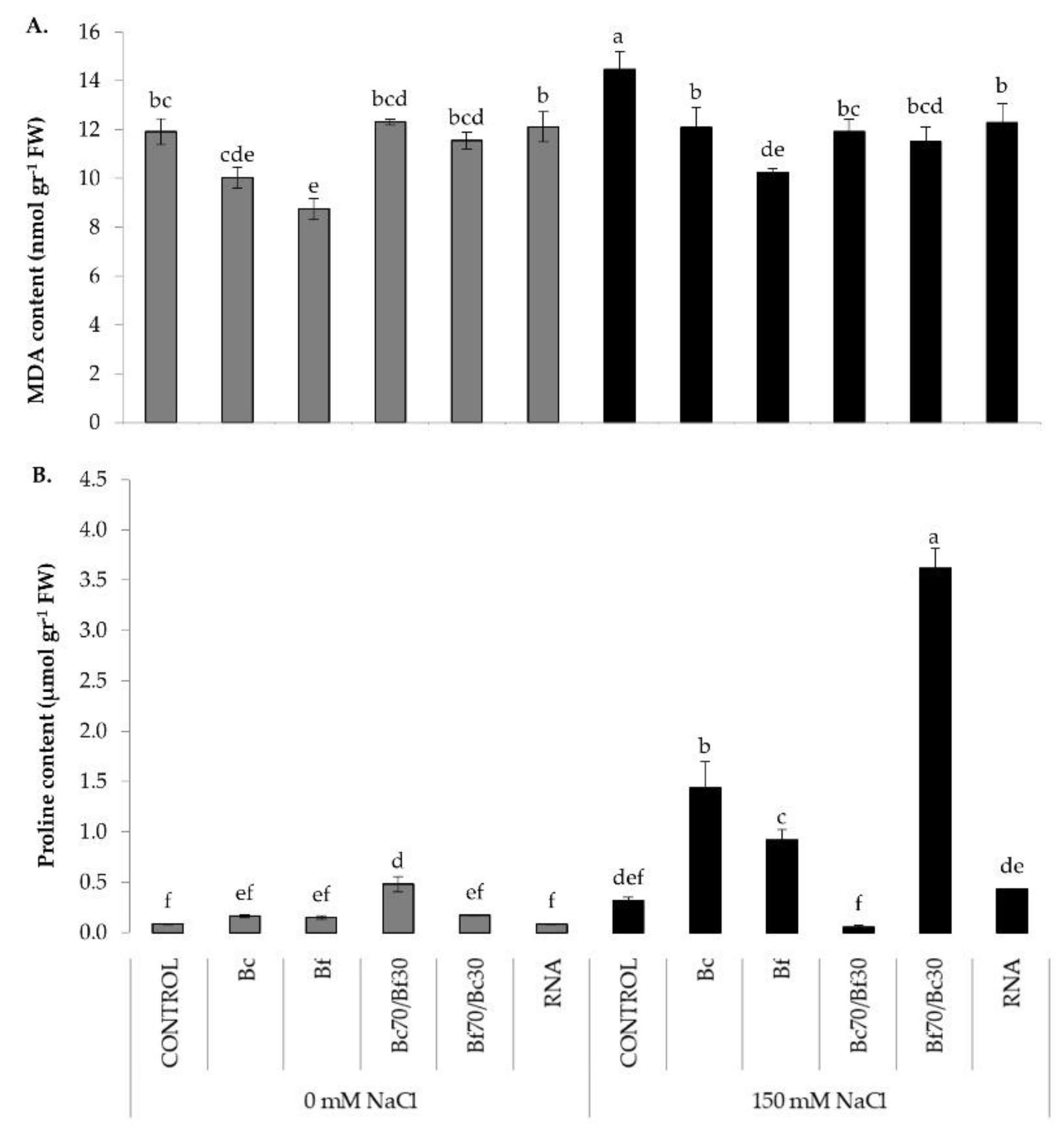
2.4. Cellular Damage and Osmoprotectant Responses of Biostimulants Treatments
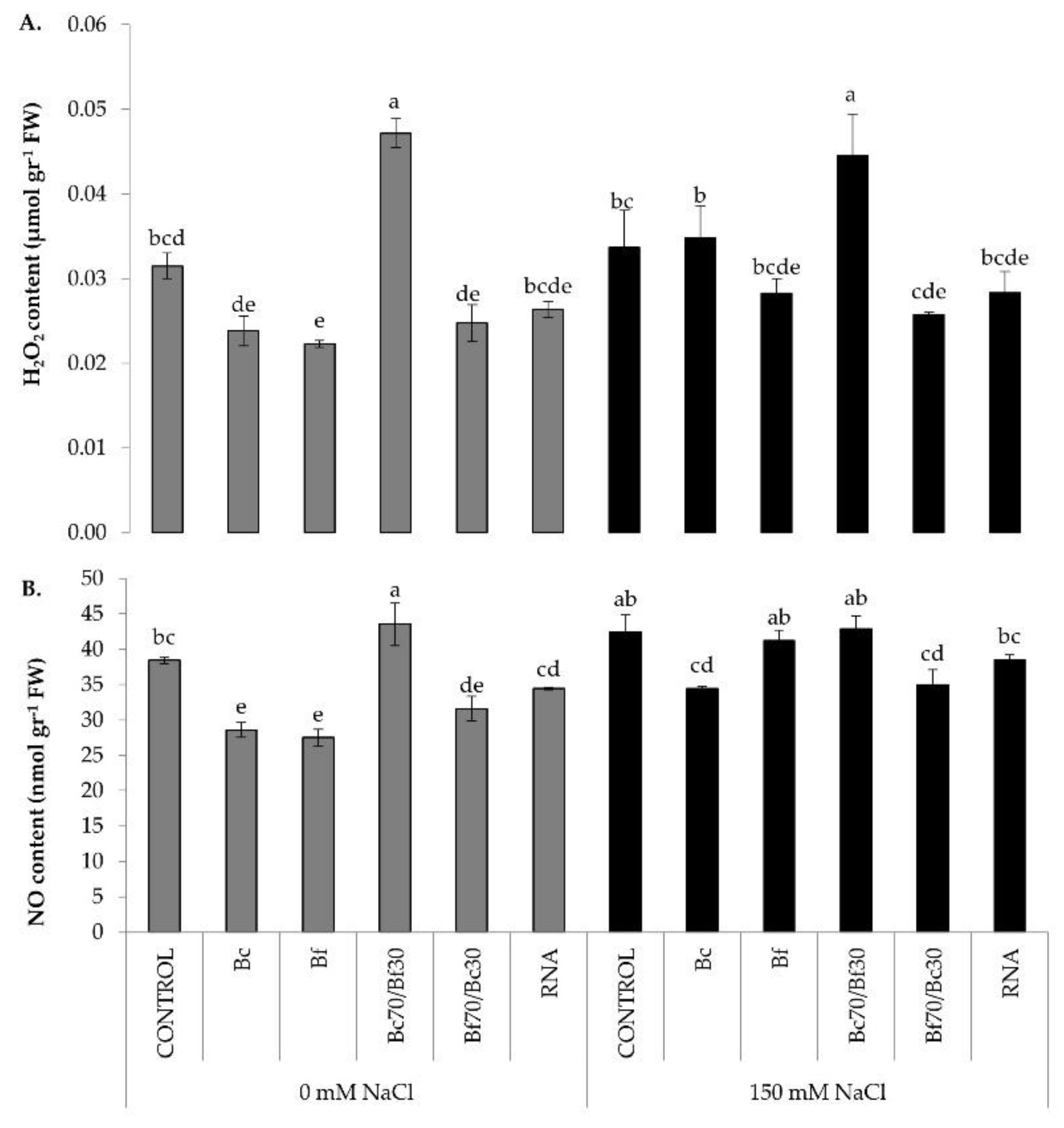
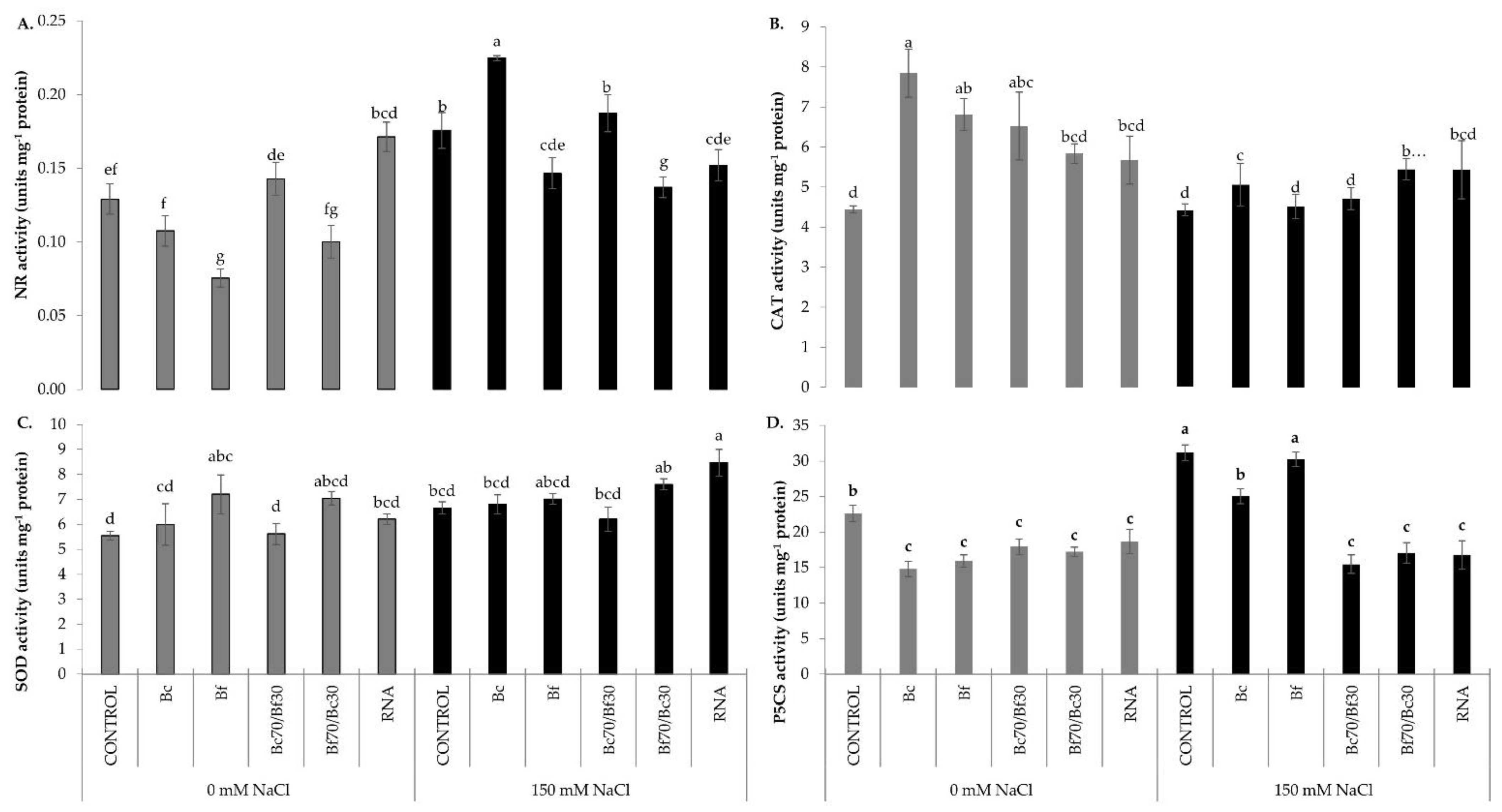
2.5. Regulation of Nitro-Oxidative Homeostasis and Osmoprotectant Biosynthesis on Biostimulant-Treated Tomatoes
2.6. Molecular Responses of Biostimulant-Treated Tomato Plants Prior to Salt Exposure
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experimental Treatments
4.2. Physiological Measurements
4.3. MDA, Reactive Species and Proline Quantification
4.4. Antioxidant Enzymatic Activities
4.5. RT-qPCR Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mariani, L.; Ferrante, A. Agronomic Management for Enhancing Plant Tolerance to Abiotic Stresses—Drought, Salinity, Hypoxia, and Lodging. Horticulturae 2017, 3, 52. [Google Scholar] [CrossRef] [Green Version]
- Coban, A.; Akhoundnejad, Y.; Dere, S.; Dasgan, H.Y. Impact of salt-tolerant rootstock on the enhancement of sensitive tomato plant responses to salinity. HortScience 2020, 55, 35–39. [Google Scholar] [CrossRef] [Green Version]
- Delian, E.; Bădulescu, L.; Dobrescu, A.; Chira, L.; Lagunovschi-Luchian, V. A brief overview of seed priming benefits in tomato. Rom. Biotechnol. Lett. 2017, 22, 12505–12513. [Google Scholar]
- Dorais, M.; Ehret, D.L.; Papadopoulos, A.P. Tomato (Solanum lycopersicum) health components: From the seed to the consumer. Phytochem. Rev. 2008, 7, 231–250. [Google Scholar] [CrossRef]
- Del Giudice, R.; Petruk, G.; Raiola, A.; Barone, A.; Monti, D.M.; Rigano, M.M. Carotenoids in fresh and processed tomato (Solanum lycopersicum) fruits protect cells from oxidative stress injury. J. Sci. Food Agric. 2016, 97, 1616–1623. [Google Scholar] [CrossRef]
- Kimura, S.; Sinha, N. Tomato (Solanum lycopersicum): A Model Fruit-Bearing Crop. Cold Spring Harb. Protoc. 2008, 2008, pdb.emo105. [Google Scholar] [CrossRef]
- Dasgan, H.Y.; Aktas, H.; Abak, K.; Cakmak, I. Determination of screening techniques to salinity tolerance in tomatoes and investigation of genotype responses. Plant Sci. 2002, 163, 695–703. [Google Scholar] [CrossRef]
- Cuartero, J.; Bolarín, M.C.; Asíns, M.J.; Moreno, V. Increasing salt tolerance in the tomato. J. Exp. Bot. 2006, 57, 1045–1058. [Google Scholar] [CrossRef]
- Hallauer, A.R. Evolution of plant breeding. Crop Breed. Appl. Biotechnol. 2011, 11, 197–206. [Google Scholar] [CrossRef] [Green Version]
- Christou, P. Plant genetic engineering and agricultural biotechnology 1983–2013. Trends Biotechnol. 2013, 31, 125–127. [Google Scholar] [CrossRef]
- Taranto, F.; Nicolia, A.; Pavan, S.; De Vita, P.; D’Agostino, N. Biotechnological and Digital Revolution for Climate-Smart Plant Breeding. Agronomy 2018, 8, 277. [Google Scholar] [CrossRef]
- Savvides, A.; Ali, S.; Tester, M.; Fotopoulos, V. Chemical Priming of Plants Against Multiple Abiotic Stresses: Mission Possible? Trends Plant Sci. 2016, 21, 329–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakur, M.; Sharma, P.; Anand, A. Seed Priming-Induced Early Vigor in Crops: An Alternate Strategy for Abiotic Stress Tolerance; Springer: Singapore, 2019. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Parvin, K.; Bardhan, K.; Nahar, K.; Anee, T.I.; Masud, A.A.C.; Fotopoulos, V. Biostimulants for the regulation of reactive oxygen species metabolism in plants under abiotic stress. Cells 2021, 10, 2537. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic Biostimulatory Action: Designing the Next Generation of Plant Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EU. Regulation of the European Parliament and of The Council Laying Down Rules on the Making Available on the Market of EU Fertilising Products and Amending Regulations (Ec) No 1069/2009 and (Ec) No 1107/2009 and Repealing Regulation (Ec) No 2003/2003. 2019. Available online: https://eur-lex.europa.eu/legalcontent/EN/TXT/?uri=OJ:L:2019:170:TOC (accessed on 16 July 2022).
- Bulgari, R.; Franzoni, G.; Ferrante, A. Biostimulants application in horticultural crops under abiotic stress conditions. Agronomy 2019, 9, 306. [Google Scholar] [CrossRef] [Green Version]
- Kocira, S.; Szparaga, A.; Hara, P.; Treder, K.; Findura, P.; Bartoš, P.; Filip, M. Biochemical and economical effect of application biostimulants containing seaweed extracts and amino acids as an element of agroecological management of bean cultivation. Sci. Rep. 2020, 10, 17759. [Google Scholar] [CrossRef]
- Du Jardin, P. Plant biostimulants: Definition, concept, main categories and regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Reynaud, H.; Canaguier, R.; Trtilek, M.; Panzarova, K.; et al. Understanding the Biostimulant Action of Vegetal-derived Protein Hydrolysates by High-throughput Plant Phenotyping and Metabolomics: A Case Study on Tomato. Front. Plant Sci. 2018, 10, 47. [Google Scholar] [CrossRef]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and crop responses: A review. Biol. Agric. Hortic. 2014, 31, 1–17. [Google Scholar] [CrossRef]
- Machado, R.; Serralheiro, R. Soil Salinity: Effect on Vegetable Crop Growth. Management Practices to Prevent and Mitigate Soil Salinization. Horticulturae 2017, 3, 30. [Google Scholar] [CrossRef] [Green Version]
- Marschner, H. Mineral Nutrition of Higher Plants, 2nd ed.; Academic Press: New York, NY, USA, 1995. [Google Scholar]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Senge, M.; Dai, Y. Effects of salinity stress on growth, yield, fruit quality and water use efficiency of tomato under hydroponics system. Rev. Agric. Sci. 2016, 4, 46–55. [Google Scholar] [CrossRef]
- Maach, M.; Boudouasar, K.; Akodad, M.; Skalli, A.; Moumen, A.; Baghour, M. Application of biostimulants improves yield and fruit quality in tomato. Int. J. Veg. Sci. 2020, 27, 288–293. [Google Scholar] [CrossRef]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural uses of plant biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef] [Green Version]
- Sharma, H.S.; Fleming, C.; Selby, C.; Rao, J.R.; Martin, T. Plant biostimulants: A review on the processing of macroalgae and use of extracts for crop management to reduce abiotic and biotic stresses. J. Appl. Phycol. 2014, 26, 465–490. [Google Scholar] [CrossRef]
- Rachidi, F.; Benhima, R.; Sbabou, L.; El Arroussi, H. Microalgae polysaccharides bio-stimulating effect on tomato plants: Growth and metabolic distribution. Biotechnol. Rep. 2020, 25, e00426. [Google Scholar] [CrossRef]
- Zou, P.; Lu, X.; Zhao, H.; Yuan, Y.; Meng, L.; Zhang, C.; Li, Y. Polysaccharides Derived from the Brown Algae Lessonia nigrescens Enhance Salt Stress Tolerance to Wheat Seedlings by Enhancing the Antioxidant System and Modulating Intracellular Ion Concentration. Front. Plant Sci. 2019, 10, 48. [Google Scholar] [CrossRef] [Green Version]
- Morcillo, R.; Manzanera, M. The Effects of Plant-Associated Bacterial Exopolysaccharides on Plant Abiotic Stress Tolerance. Metabolites 2021, 11, 337. [Google Scholar] [CrossRef]
- Türkmen, Ö.; Dursun, A.; Turan, M.; Erdinç, Ç. Calcium and humic acid affect seed germination, growth, and nutrient content of tomato (Lycopersicon esculentum L.) seedlings under saline soil conditions. Acta Agric. Scand. Sect. B Soil Plant Sci. 2004, 54, 168–174. [Google Scholar] [CrossRef]
- Campobenedetto, C.; Mannino, G.; Beekwilder, J.; Contartese, V.; Karlova, R.; Bertea, C.M. The application of a biostimulant based on tannins affects root architecture and improves tolerance to salinity in tomato plants. Sci. Rep. 2021, 11, 354. [Google Scholar] [CrossRef] [PubMed]
- Volpe, V.; Chitarra, W.; Cascone, P.; Volpe, M.G.; Bartolini, P.; Moneti, G.; Pieraccini, G.; Di Serio, C.; Maserti, B.; Guerrieri, E.; et al. The Association with Two Different Arbuscular Mycorrhizal Fungi Differently Affects Water Stress Tolerance in Tomato. Front. Plant Sci. 2018, 9, 1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sena, J.O.A.; Zaidan, H.A.; Castro, P.R.d.C.e. Transpiration and stomatal resistance variations of perennial tropical crops under soil water availability conditions and water deficit. Braz. Arch. Biol. Technol. 2007, 50, 225–230. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef] [Green Version]
- Kałużewicz, A.; Krzesiński, W.; Spiżewski, T.; Zaworska, A. Effect of Biostimulants on Several Physiological Characteristics and Chlorophyll Content in Broccoli under Drought Stress and Re-watering. Not. Bot. Horti Agrobot. Cluj-Napoca 2017, 45, 197. [Google Scholar] [CrossRef] [Green Version]
- Seciu, A.-M.; Oancea, A.; Gaspar, A.; Moldovan, L.; Craciunescu, O.; Stefan, L.; Petrus, V.; Georgescu, F. Water Use Efficiency on Cabbage and Cauliflower Treated with a New Biostimulant Composition. Agric. Agric. Sci. Proc. 2016, 10, 475–484. [Google Scholar] [CrossRef] [Green Version]
- Mäkelä, P.; Munns, R.; Colmer, T.D.; Condon, A.G.; Peltonen-Sainio, P. Effect of foliar applications of glycine betaine on stomatal conductance, abscisic acid and solute concentrations in leaves of salt- or drought-stressed tomato. Aust. J. Plant Physiol. 1998, 25, 655. [Google Scholar] [CrossRef]
- Gharbi, F.; Zribi, L.; Daly, A.; Rejeb, S.; Hanchi, B. Photosynthetic Responses of Tomato Leaves to Salt and Cadmium Stresses: Growth and Chlorophyll a Fluorescence Kinetic Analyses. Pol. J. Environ. Stud. 2018, 27, 2499–2508. [Google Scholar] [CrossRef]
- Peripolli, M.; Dornelles, S.; Lopes, S.; Tabaldi, L.; Trivisiol, V. Application of biostimulants in tomato subjected to water deficit: Physiological, enzymatic and production responses. Rev. Bras. Engenharia Agrícola Ambient. 2021, 25, 90–95. [Google Scholar] [CrossRef]
- Kong, W.; Liu, F.; Zhang, C.; Zhang, J.; Feng, H. Non-destructive determination of Malondialdehyde (MDA) distribution in oilseed rape leaves by laboratory scale NIR hyperspectral imaging. Sci. Rep. 2016, 6, 35393. [Google Scholar] [CrossRef]
- Turan, M.; Yildirim, E.; Ekinci, M.; Argin, S. Effect of Biostimulants on Yield and Quality of Cherry Tomatoes Grown in Fertile and Stressed Soils. HortSci. Horts 2021, 56, 414–423. [Google Scholar] [CrossRef]
- Rady, M.M.; Taha, R.S.; Mahdi, A.H.A. Proline enhances growth, productivity and anatomy of two varieties of Lupinus termis L. grown under salt stress. S. Afr. J. Bot. 2016, 102, 221–227. [Google Scholar] [CrossRef]
- Alfosea-Simón, M.; Zavala-Gonzalez, E.A.; Camara-Zapata, J.M.; Martínez-Nicolás, J.J.; Simón, I.; Simón-Grao, S.; García-Sánchez, F. Effect of foliar application of amino acids on the salinity tolerance of tomato plants cultivated under hydroponic system. Sci. Hortic. 2020, 272, 109509. [Google Scholar] [CrossRef]
- Foyer, C.; Noctor, G. Redox sensing and signalling associated with reactive oxygen in chloroplasts, peroxisomes and mitochondria. Physiol. Plant 2003, 119, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Gohari, G.; Panahirad, S.; Sepehri, N.; Akbari, A.; Zahedi, S.M.; Jafari, H.; Dadpour, M.R.; Fotopoulos, V. Enhanced tolerance to salinity stress in grapevine plants through application of carbon quantum dots functionalized by proline. Environ. Sci. Pollut. Res. 2021, 28, 42877–42890. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2017, 54, 287–293. [Google Scholar] [CrossRef] [Green Version]
- Abogadallah, G.M. Antioxidative defense under salt stress. Plant Signal. Behav. 2010, 5, 369–374. [Google Scholar] [CrossRef] [Green Version]
- Bano, S.; Ashraf, M.; Akram, N.A.; Al-Qurainy, F. Regulation in some vital physiological attributes and antioxidative defense system in carrot (Daucus carota L.) under saline stress. J. Appl. Bot. Food Qual. 2012, 85, 105–115. [Google Scholar]
- Wendehenne, D.; Durner, J.; Klessig, D.F. Nitric oxide: A new player in plant signalling and defence responses. Curr. Opin. Plant Biol. 2004, 7, 449–455. [Google Scholar] [CrossRef]
- Neill, S.; Desikan, R.; Hancock, J. Hydrogen peroxide signalling. Curr. Opin. Plant Biol. 2002, 5, 388–395. [Google Scholar] [CrossRef]
- Beevers, L.; Hageman, R.H. Nitrate Reduction in Higher Plants. Ann. Rev. Plant Physiol. 1969, 20, 495–522. [Google Scholar] [CrossRef]
- Lane, H.C.; Thompson, A.C.; Hesketh, J.D.; Sloane, C. Some observations on nitrate reduction in cotton. In Proceedings of the Beltwide Cotton Production Research Conferences; Brown, J., Ed.; National Cotton Council: Memphis, TN, USA, 1975. [Google Scholar]
- Flores, P.; Navarro, J.M.; Garrido, C.; Ribio, J.S.; Martınez, V. Influence of Ca2+, K+ and NO3− fertilisation on nutritional quality of pepper. J. Sci. Food Agric. 2004, 84, 569–574. [Google Scholar] [CrossRef]
- Filippou, P.; Antoniou, C.; Yelamanchili, S.; Fotopoulos, V. NO loading: Efficiency assessment of five commonly used application methods of sodium nitroprusside in Medicago truncatula plants. Plant Physiol. Biochem. 2012, 60, 115–118. [Google Scholar] [CrossRef] [PubMed]
- Ertani, A.; Schiavon, M.; Nardi, S. Transcriptome-Wide Identification of Differentially Expressed Genes in Solanum lycopersicon L. in Response to an Alfalfa-Protein Hydrolysate Using Microarrays. Front. Plant Sci. 2017, 8, 1159. [Google Scholar] [CrossRef] [Green Version]
- Campobenedetto, C.; Mannino, G.; Agliassa, C.; Acquadro, A.; Contartese, V.; Garabello, C.; Bertea, C.M. Transcriptome Analyses and Antioxidant Activity Profiling Reveal the Role of a Lignin-Derived Biostimulant Seed Treatment in Enhancing Heat Stress Tolerance in Soybean. Plants 2020, 9, 1308. [Google Scholar] [CrossRef] [PubMed]
- Huot, B.; Yao, J.; Montgomery, B.L.; He, S.Y. Growth—Defense Tradeoffs in Plants: A Balancing Act to Optimize Fitness. Mol. Plant 2014, 7, 1267–1287. [Google Scholar] [CrossRef] [Green Version]
- Christou, A.; Manganaris, G.A.; Papadopoulos, I.; Fotopoulos, V. Hydrogen sulfide induces systemic tolerance to salinity and non-ionic osmotic stress in strawberry plants through modification of reactive species biosynthesis and transcriptional regulation of multiple defense pathways. J. Exp. Bot. 2013, 64, 1953–1966. [Google Scholar] [CrossRef]
- Loreto, F.; Velikova, V. Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quenches ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiol. 2001, 127, 1781–1787. [Google Scholar] [CrossRef] [PubMed]
- Filippou, P.; Bouchagier, P.; Skotti, E.; Fotopoulos, V. Proline and reactive oxygen/nitrogen species metabolism is involved in the tolerant response of the invasive plant species Ailanthus altissima to drought and salinity. Environ. Exper. Bot. 2014, 97, 1–10. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Yang, S.-L.; Lan, S.-S.; Gong, M. Hydrogen peroxide-induced proline and metabolic pathway of its accumulation in maize seedlings. J. Plant Physiol. 2009, 166, 1694–1699. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expressions software tool (REST(C)) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, 2002–2036. [Google Scholar] [CrossRef] [PubMed]
- Lovdal, T.; Lillo, C. Reference gene selection for quantitative real-time PCR normalization in tomato subjected to nitrogen, cold, and light stress. Anal. Biochem. 2009, 387, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Martinez, V.; Nieves-Cordones, M.; Lopez-Delacalle, M.; Rodenas, R.; Mestre, T.; Garcia-Sanchez, F.; Rubio, F.; Nortes, P.A.; Mittler, R.; Rivero, R.M. Tolerance to Stress Combination in Tomato Plants: New Insights in the Protective Role of Melatonin. Molecules 2018, 23, 535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asins, M.J.; Villalta, I.; Aly, M.M.; Olías, R.; Alvarez de Morales, P.; Huertas, R.; Belver, A. Two closely linked tomato HKT coding genes are positional candidates for the major tomato QTL involved in Na+/K+ ho-meostasis. Plant Cell Environ. 2013, 36, 1171–1191. [Google Scholar]
- Almutairi, Z. Effect of nano-silicon application on the expression of salt tolerance genes in germinating tomato (Solanum lycopersicum L.) seedlings under salt stress. Plant Omics J. 2016, 9, 106–114. [Google Scholar]
- Huang, S.; Gao, Y.; Liu, J.; Peng, X.; Niu, X.; Fei, Z.; Cao, S.; Liu, Y. Genome-wide analysis of WRKY transcription factors in Solanum lycopersicum. Mol. Genet. Genom. 2012, 287, 495–513. [Google Scholar]
- Lopez-Delacalle, M.; Camejo, D.M.; García-Martí, M.; Nortes, P.A.; Nieves-Cordones, M.; Martínez, V.; Rubio, F.; Mittler, R.; Rivero, R.M. Genome-wide analysis of WRKY transcription factors in Solanum lycopersicum. Front. Plant Sci. 2020, 10, 1702. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Height (cm) | Number of Leaves | Stem Width (cm) | 3rd Leaf FW (g) | Plant FW (g) | Plant DW (g) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | ||
| 0 mM NaCl | CONTROL | 14.70a | ±0.37 | 8.83a | ±0.27 | 0.514e | ±0.014 | 2.00a,b,c | ±0.19 | 8.71c,d | ±0.58 | 1.20c,d | ±0.07 |
| Bc | 14.93a | ±0.33 | 8.58a | ±0.15 | 0.597a,b | ±0.011 | 2.32a | ±0.15 | 11.34b | ±0.80 | 1.45a,b | ±0.09 | |
| Bf | 14.57a,b | ±0.23 | 8.50a | ±0.15 | 0.605a,b | ±0.013 | 2.33a | ±0.14 | 11.62a,b | ±0.47 | 1.50a | ±0.07 | |
| Bc70/Bf30 | 13.46c,d,e | ±0.37 | 7.83b,c | ±0.11 | 0.516e | ±0.009 | 1.53d,e | ±0.07 | 6.83e | ±0.22 | 1.28b,c,d | ±0.10 | |
| Bf70/Bc30 | 15.18a | ±0.60 | 8.75a | ±0.25 | 0.633a | ±0.016 | 1.90b,c,d | ±0.16 | 12.79a | ±0.41 | 1.40a,b,c | ±0.09 | |
| RNA | 14.80a | ±0.38 | 8.58a | ±0.19 | 0.534d,e | ±0.012 | 1.98a,b | ±0.12 | 9.30c,d | ±0.65 | 1.31a,b,c,d | ±0.09 | |
| 150 mM NaCl | CONTROL | 13.45c,d,e | ±0.34 | 8.33a,b | ±0.14 | 0.523d,e | ±0.009 | 1.76b,c,d | ±0.14 | 8.17d,e | ±0.46 | 1.13d | ±0.05 |
| Bc | 13.63b,c,d | ±0.24 | 8.75a | ±0.14 | 0.558c,d | ±0.012 | 2.08a,b | ±0.10 | 9.34c,d | ±0.38 | 1.23b,c,d | ±0.04 | |
| Bf | 12.83c,d | ±0.31 | 8.50a | ±0.15 | 0.517e | ±0.012 | 1.83b,c,d | ±0.08 | 8.69c,d | ±0.41 | 1.21c,d | ±0.05 | |
| Bc70/Bf30 | 12.37d | ±0.27 | 7.50c | ±0.15 | 0.468f | ±0.017 | 1.31e | ±0.05 | 5.99f | ±0.32 | 0.83e | ±0.03 | |
| Bf70/Bc30 | 14.41a,b,c | ±0.34 | 8.67a | ±0.26 | 0.577b,c | ±0.018 | 1.69c,d | ±0.10 | 10.22b,c | ±0.35 | 1.35a,b,c,d | ±0.04 | |
| RNA | 12.98c,d | ±0.29 | 8.25a,b | ±0.13 | 0.518d,e | ±0.011 | 1.92a,b,c | ±0.13 | 8.13d,e | ±0.41 | 1.14d | ±0.05 | |
| Fold Change/Treatment | C/150 VS C/0 | Bc/150 VS Bc/0 | Bf/150 VS Bf/0 | Bc70/Bf30/150 VS Bc70/Bf30/0 | Bf70/Bc30/150 VS Bf70/Bc30/0 | RNA/150 VS RNA/0 | |
|---|---|---|---|---|---|---|---|
| Gene | |||||||
| Antioxidant and other defense-related genes | SlCu/Zn-SOD | 4.77 ** | 1.22 | −1.78 | 2.80 * | 1.31 | 1.56 |
| SlFe-SOD | −1.45 | 4.86 ** | −2.50 ** | −1.04 | −3.18 ** | −1.19 | |
| SlCAT1 | 1.06 | −1.40 | −1.23 | −1.20 | −2.10 ** | 1.04 | |
| SlcAPX | 1.37 | −1.34 | −1.86 * | 2.80 * | −2.33 ** | −1.14 | |
| LOX1 | 1.73 | 2.03 * | −1.96 | 1.04 | −1.09 | 2.61 * | |
| Nitrogen and proline metabolism-related genes | P5CS | 1.51 | −3.71 * | −1.87 | −1.02 | 1.41 | 1.91 * |
| SlNR | −1.98 ** | 2.04 | −1.48 | 2.52 * | 1.18 | 1.07 | |
| SlNiR | −1.69 ** | 5.22 * | −1.20 | 1.09 | −1.06 | 1.59 | |
| Transporters | HKT1.1 | 1.69 | 2.21 * | −1.39 | 1.50 | −1.08 | 1.30 * |
| HKT1.2 | 1.30 | 1.05 | −1.77 | 1.34 * | −2.83 * | 2.60 * | |
| SlGTS1 | −1.38 | 1.26 | −1.19 | 2.26 * | −2.14 * | −1.05 | |
| Transcription factors | SlWRKY 8 | 1.19 | 1.24 | 1.27 | 2.15 * | −1.68 | 1.53 |
| SlWRKY 31 | 1.18 | 1.99 * | −1.85 * | 2.39 * | −2.97 ** | 1.23 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gedeon, S.; Ioannou, A.; Balestrini, R.; Fotopoulos, V.; Antoniou, C. Application of Biostimulants in Tomato Plants (Solanum lycopersicum) to Enhance Plant Growth and Salt Stress Tolerance. Plants 2022, 11, 3082. https://doi.org/10.3390/plants11223082
Gedeon S, Ioannou A, Balestrini R, Fotopoulos V, Antoniou C. Application of Biostimulants in Tomato Plants (Solanum lycopersicum) to Enhance Plant Growth and Salt Stress Tolerance. Plants. 2022; 11(22):3082. https://doi.org/10.3390/plants11223082
Chicago/Turabian StyleGedeon, Stella, Andreas Ioannou, Raffaella Balestrini, Vasileios Fotopoulos, and Chrystalla Antoniou. 2022. "Application of Biostimulants in Tomato Plants (Solanum lycopersicum) to Enhance Plant Growth and Salt Stress Tolerance" Plants 11, no. 22: 3082. https://doi.org/10.3390/plants11223082