
Biocontrol Activity of Aromatic and Medicinal Plants and Their Bioactive Components against Soil-Borne Pathogens

1
Department of Food Science, Albert Casimir Faculty at Mosonmagyarovar, Szechenyi Istvan University, 15-17 Lucsony Street, 9200 Mosonmagyarovar, Hungary
2
Wittmann Antal Multidisciplinary Doctoral School in Plant, Animal, and Food Sciences, Szechenyi Istvan University, 2 Var Square, 9200 Mosonmagyarovar, Hungary
3
Kisalfoldi Agricultural Ltd., 1 Fo Street, 9072 Nagyszentjanos, Hungary
*
Author to whom correspondence should be addressed.
Plants 2023, 12(4), 706; https://doi.org/10.3390/plants12040706
Submission received: 31 December 2022
/
Revised: 30 January 2023
/
Accepted: 2 February 2023
/
Published: 5 February 2023
(This article belongs to the Special Issue On the Microbe-Essential Oil Interplay: Processes, Mechanisms, Impacts and Applications)
Abstract
:Soil-borne phytopathogens can have detrimental effects on both cereal and horticultural crops resulting in serious losses worldwide. Due to their high efficiency and easy applicability, synthetic pesticides are still the primary choice in modern plant disease control systems, but stringent regulations and increasing environmental concerns make the search for sustainable alternatives more pressing than ever. In addition to the incorporation of botanicals into agricultural practices, the diversification of cropping systems with aromatic and medicinal plants is also an effective tool to control plant diseases through providing nutrients and shaping soil microbial communities. However, these techniques are not universally accepted and may negatively affect soil fertility if their application is not thoroughly controlled. Because the biocontrol potential of aromatic and medicinal plants has been extensively examined over the past decades, the present study aims to overview the recent literature concerning the biopesticide effect of secondary metabolites derived from aromatic and medicinal plants on important soil-borne plant pathogens including bacteria, fungi, and nematodes. Most of the investigated herbs belong to the family of Lamiaceae (e.g., Origanum spp., Salvia spp., Thymus spp., Mentha spp., etc.) and have been associated with potent antimicrobial activity, primarily due to their chemical constituents. The most frequently tested organisms include fungi, such as Rhizoctonia spp., Fusarium spp., and Phytophthora spp., which may be highly persistent in soil. Despite the intense research efforts dedicated to the development of plant-based pesticides, only a few species of aromatic herbs are utilized for the production of commercial formulations due to inconsistent efficiency, lack of field verification, costs, and prolonged authorization requirements. However, recycling the wastes from aromatic and medicinal plant-utilizing industries may offer an economically feasible way to improve soil health and reduce environmental burdens at the same time. Overall, this review provides comprehensive knowledge on the efficiency of aromatic herb-based plant protection techniques, and it also highlights the importance of exploiting the residues generated by aromatic plant-utilizing sectors as part of agro-industrial processes.
1. Introduction
Continuously increasing human population, ever-growing demand for food, and agricultural intensification have been placing a tremendous pressure on soil health, leading to degradation and exhaustion of agricultural lands and declining yields and productivity [1,2,3,4]. The evaluation of soil health and quality is a critically important step in maintaining long-term agricultural sustainability [5,6]. Healthy soil is characterized by stability, continuous nutrient cycling, stress resilience, richness, evenness, abundance, and diversity of microbial communities [7,8]. The interactions between plants and soil-dwelling microorganisms are important drivers of the agroecosystem, as these indigenous microbes can control the development, defense, tolerance, and nutrition of plants [9,10,11]. However, the composition and abundance of microbial communities are highly influenced by various parameters such as the type of cultivated plants, environmental fluctuations, and conducted agricultural activities [7,10,12].
Reduced plant productivity and phytopathogen-related disease outbreaks can be considered as indicators of soil instability, poor ecosystem health, and a lack of microbial diversity [7,8]. Soil-borne pathogens including viruses, bacteria, fungi, protozoa, and nematodes are well known for their socio-economic and ecological impacts on the ecosystem by causing serious losses in the agro-industry [13,14,15,16]. Plant roots and their exudates provide substrates and space for these phytopathogens leading to colonization and infection. Generally, the eradication of these organisms is especially hard in continuous-cropping systems, due to the intensive tillage and cultivation processes [7,17].
Both agricultural practices and directed agronomical measures can affect the diversity of soil microbial communities [18]. Traditionally, crop rotation techniques had been used by farmers to manage the productivity of soil and reduce the possibility of crop diseases [19]; however, after World War II, shortened rotation periods, intensive tillage, monocropping, and application of synthetic pesticides and fertilizers came to the fore [20]. Although numerous cultural, physical, biological, and genetic options are available nowadays, the main strategies for controlling soil-borne pathogens are still highly dependent on the use of agrochemicals [15,21] due to their high efficiency and easy applicability [22]. Besides the positive effects, the indiscriminate use of these chemical agents may induce undesirable changes to the beneficial soil microorganisms and human and animal health [22,23,24]. The commonly applied chemical pesticides are usually effective against their target organisms, but they are not specific for them [25]. According to Ashraf and Zuhaib [26], only an estimated 0.1% of the used synthetic chemicals reach the targeted pathogens, whereas the remaining 99.9% may contaminate the environment and endanger nontarget organisms.
The application of many chemical agents has been restricted in certain parts of the world [27]. Therefore, finding sustainable and safer solutions with minimum or no side effects is essential to develop effective soil health management strategies and lower the possible environmental pollution [23,28,29,30].
In recent years, there has been an increasing interest in the application of allelochemicals isolated from higher plants in plant protection because they are safer than their synthetic counterparts, effective at reduced doses, and biodegradable without leaving harmful residues [31,32,33,34]. The use of pathogen-suppressing secondary metabolites (e.g., alkaloids, terpenoids, phenols) is especially important in organic farming systems [33,35], but conventional cultivation methods and agricultural practices utilizing aromatic plants and their residues (i.e., solid biomass and hydrosols) may also be effective in controlling the spread and severity of plant diseases (Figure 1).
This paper summarizes and reviews (a) the most important soil-borne pathogens including bacteria, fungi, nematodes, and viruses; (b) the current status of botanical pesticides; and (c) the beneficial effects of aromatic and medicinal plant-based cultivation techniques on soil and plant health. Moreover, the reutilization of by-products generated by aromatic plant-based industries as biocontrol agents is also discussed.
2. Phytopathogens in Soil
Soil biology is in direct connection with agricultural sustainability [36], as healthy soil can recycle nutrients, decompose organic matter, support biological activities, suppress the growth of pathogens, and inactivate toxic compounds [37]. It is the habitat of a myriad of organisms (e.g., invertebrate animals, plants, protozoa, fungi, and bacteria) that play an important role in maintaining agro-ecosystem functions and productivity [38,39]. As for microbial diversity, only a few grams of soil can contain millions of species [40]. In addition to beneficial microbes, however, major and minor pathogens may also be present [15,41]. Even though the occurrence of microbial plant diseases is a natural part of crop production, the excessive accumulation and epidemic spread of phytopathogens can induce serious crop damages and yield losses [12,36].
Soil-borne pathogenic fungi, bacteria, protozoa, viruses, and plant parasitic nematodes can cause negative plant–soil feedback individually or collectively [42,43]. They usually affect the root system or the stem base of plants, but vascular diseases caused by these organisms have also been reported [44]. Overall, approximately 15% of total crop production is estimated to be lost to different biological threats annually [45], but microbe-induced diseases may even lead to total crop failure [46].
2.1. Fungi
As far as fungal pathogens are concerned, infections can be transmitted through soil-damaging agricultural and horticultural products [47]. Fusarium, Verticillium, Rhizoctonia, Sclerotinia, Phytophthora, and Pythium species may survive in the soil and in plant residues for longer periods by forming resistant structures such as sclerotia, microsclerotia, oospores, chlamydospores, or hyphae [22,48]. Several of them can invade the host plant through roots and stems or spread rapidly among the seedlings [49] causing root necrosis and vascular diseases, as well as rot, gall, and proliferation of roots and tubers [44]. Recent studies have highlighted that the causal agents of root rot (Fusarium solani and Fusarium oxysporum f. sp. dahliae-lycopersici) and wilting (Verticillium dahliae) are able to infect a considerable range of crop plants, making the controlling process extremely difficult without high-cost fumigants [50]. For instance, Verticillium dahliae can survive in the soil for up to 14 years by forming microsclerotia and may affect more than 160 plant species [51]. Furthermore, fungi belonging to the genera Pythium and Phytophthora may cause similar symptoms in the crown, stem, and root tissues of certain crops leading to scars; girdled stem; stunt; stem lesions; foliar blight; browning; wilting; decay; damp-off; loss of root density; and rot of crown, roots, and fruits [52,53].
2.2. Bacteria
Along with fungal strains, important bacterial pathogens can be found in the soil including Agrobacterium, Pectobacterium, Pseudomonas, Ralstonia, and Xanthomonas species [36,54]; however, bacterial pathogens are less likely to cause plant diseases, as they need a wound or a natural opening to invade the host plant [16]. Ralstonia solanacearum is a causal agent of bacterial wilt disease in various crops, including potato, banana, tomato, and peanut. The bacteria enter the plant through wounds, cracks, or root tips; trigger morphological alterations in the root system of the infected host; invade the xylem vessels; and cause wilting symptoms and plant death [54,55]. Pseudomonas syringae and Pectobacterium carotovorum are also important soil-borne bacteria inducing necrotic lesions on tomato and carrot soft root, respectively [56]. Pectobacterium atrosepticum is mainly responsible for various field symptoms of potato, including reduced emergence, wilting, chlorosis, stem and tuber rot, haulm desiccation, blackleg, and plant death [57]. Xanthomonas campestris pv. musacearum may infect banana plants through injured roots and stems. Since no remedy is available against this pathogen, the infected plant may be cut down, and fallowing or prolonged crop rotation technique may be introduced [58].
2.3. Nematodes
So far, over 4100 species of plant-parasitic nematodes have been identified. They cause serious economic losses globally (approximately USD 173 billion a year [59]) due to their wide variety of interactions and host range. These small roundworms can be categorized as ectoparasitic and endoparasitic species [59,60,61,62,63,64,65,66]. Among them, the sedentary endoparasitic nematodes [66] are the most economically important species. The temperate Meloidogyne hapla and the tropical Meloidogyne incognita, Meloidogyne arenaria, and Meloidogyne javanica belong to the most destructive root-knot nematodes that can infect almost all vascular plant species [61]. The parasitism of these biotrophic organisms includes the establishment of permanent feeding sites in the root cortex, vascular parenchyma, endodermis, and pericycle of host plants. The intensified metabolic activity in the multinucleate giant cells mobilizes photosynthates and, consequently, decreases plant quality and quantity with typical symptoms such as suppressed plant growth, wilting, leaf discoloration, and root deformation [61,67]. Southern knot-nematodes may also interact with soil-borne fungi resulting in disease complexes [62]. Regarding cyst nematodes, potato, cereal, and soybean cyst nematodes have been identified as the most significant obligate biotrophs that may be highly persistent in the soil and can survive for longer periods as eggs inside a cyst without a host making their eradication almost impossible [61,68]. In addition, ectoparasitic nematodes are an integral part of the soil fauna [69] moving freely between root-feeding sites. However, they are more likely exposed to various environmental conditions and predators. Similarly to endoparasites, they cause localized damage weakening the host plant defense against bacterial and fungal infections [70], whereas certain species, including Xiphinema index, may act as a vector of viruses [63]. Currently, various methods (e.g., use of chemical agents, bio-fumigants, sanitation, resistant species, solarization, steaming, tilling, fallowing, escape cropping, cover cropping with trap crops, crop rotation with poor or non-hosts, soil amendments, biological control by natural antagonists, etc.) are employed to reduce nematode infestation in the soil [71]. Although chemical nematicides are efficient for the control of root-knot nematodes, their use has been gradually decreased due to their toxicity, limited availability in developing countries, high costs, and impaired efficacy after repeated applications [72,73].
2.4. Viruses
Soil may contain enormous amounts of soil-borne plant viruses with an estimated viral abundance of 1031 [74]. The annual crop losses caused by these tiny organisms are estimated at USD 60 billion worldwide. Currently, more than 2000 species have been identified that affect economically important crops including barley, groundnut, wheat, sugar beet, potato, and fruit crops [42,75]. The first report was released in 1925 in connection with a soil-borne virus, the soil-borne wheat mosaic virus, and further 53 plant pathogens belonging to 12 genera have been recognized as soil-borne viruses in the last decades [27]. They are generally transmitted via soil, fungi, nematodes, plasmodiophora, insects and other arthropods, sap transmission, seed and pollen transmission, mechanical friction, and vegetative propagation materials [42,74]. In general, they are extremely persistent [42] just as with beet soil-borne mosaic virus that is spread by a soil-borne plasmodiophoromycete, Polymyxa betae, which generates resting spores allowing the virus to survive in the soil for decades [76]. Although different viruses have different host ranges, collectively, they can infect almost all kinds of cultivated and noncultivated plants, and the infections can be transmitted from the affected plants to the healthy ones [75]. As an example, the tobacco mosaic virus is a common viral disease that affects over 1000 species in 85 plant families [75,77]. The remains of infected plants in the soil are the principal reservoirs of the virus. Transmission mostly occurs when leaves are rubbed in the presence of virus-containing soil or when injured root hairs come into contact with infected residues or free virus [78]. Furthermore, tomato spotted wilt virus is capable of infecting more than 1000 plant species (e.g., ornamental plants, lettuce, pepper, potato, etc.) [79]. Since the limitation of viral infections and the eradication of viruses from infected soil are almost impossible, control methods have mostly relied on various agrochemicals that can reduce the vector population, resistant plant varieties, and other preventive measures [27,42,80].
3. Non-Chemical Plant Protection Methods
The lack of effective pest control is still a major global obstacle to improved plant health and productivity [81]. Synthetic agrochemicals have been widely used to control soil-borne pathogens [21], but the regulations on the application of these products have become more stringent because of their adverse effects [82]. After the implementation of the Montreal Protocol, EU regulations EC 2037/2000 and EC 3093/1994 completely prohibited the use of methyl bromide, the most popular fumigant against soil-borne bacteria, fungi, and nematodes. Furthermore, European Directive 2009/128/EC on the sustainable use of pesticides was issued to reduce the risk of pesticides to the environment and human health [22,28,83,84,85], and European Regulation no. 1107/2009 has been controlling the placement of plant protection products on the market [86]. This led to the prohibition of several traditional pesticides [87] and fueled the search for other suitable candidates, including existing and new agrochemicals and more sustainable, economically feasible alternatives that are less dangerous to the environment [88,89]. However, these methods must meet various standards regarding pest specificity, toxicity, pesticide resistance, cost, and availability [90].
Organic farming is a rapidly developing more sustainable method that improves soil quality [38,91,92], thereby lowering the risk of negative environmental impacts such as ecosystem degradation and global warming [93]. This form of agricultural production aims to utilize non-chemical disease prevention tools rather than other treatments [92]. For instance, ecofriendly farming systems rely on the use of different physical, cultural, and biological management techniques to suppress the spread of pathogenic microorganisms [94]. These methods, with only a few exceptions, do not pollute the environment [95]. Nevertheless, the activity spectrum of non-chemical farming strategies is extremely variable, and they are often less effective than their chemical analogues [96,97]. Therefore, successful disease control may require the implementation of multiple methods [98]. Yoon et al. [79] used soil-dwelling predatory mites and an essential oil mixture to control tomato spotted wilt virus and thrips. Likewise, Schmitt and Seddon [99] reported that the simultaneous use of microbiological control agents and plant extracts may have an additive or even a synergistic effect. Meanwhile, Baysal-Gurel et al. [48] showed that cover crops in combination with solarization significantly reduced Rhizoctonia root rot severity.
As another option, Integrated Pest Management (IPM) is a redesigned form of intensive agricultural systems, in which non-chemical methods, botanicals, or other curative treatments are combined with synthetic chemicals [35]. Just as with organic production, the goal is not to fully eliminate disease-causing organisms but to prevent them from becoming damaging or dominant in the cropping system. As a result, an integration of different preventive tactics (i.e., chemical, biological, physical, and genetic means with cropping practices) may be more effective than a single method [100,101]. A survey conducted by Kabir and Rainis [102] showed that vegetable farmers in Bangladesh used additional methods (e.g., biological control, soil amendments, pheromone traps, manual cleaning, soil solarization, and botanicals), while pure pesticides were only applied when there were no other options, or the pest infestation was high. In a study by Fielding et al. [103] eight medicinal plant extracts were applied successfully in combination with kresoxim-methyl for the inhibition of gray mold on apples. Shlevin et al. [104] analyzed independent studies to evaluate the efficacy of soil solarization with fumigants in controlling various soil-borne pathogens (e.g., Fusarium species; root-knot nematodes; and the group of Sclerotium cepivorum, Verticillium spp., Pyrenochaeta spp., Rhizoctonia spp., and Pythium spp.). Overall, their results showed that the combined technique improved management efficacy compared to soil solarization only. In contrast, Deguine et al. [101] reported that these integrated methods were rarely adopted by farmers, and, as a result, pesticides remained the cornerstone of crop protection around the world.
4. Aromatic Plants as Biocontrol Agents
There are approximately 17,500 aromatic plant species throughout the world, either cultivated or collected in the wild, that are greatly used for cosmetic, preservation, flavoring, therapeutic, and pharmacologic purposes [105,106,107,108]. These herbs are responsible for the synthesis of a multitude of secondary metabolites (e.g., flavonoids, phenol compounds, terpenes, and nitrogen-containing chemicals), which are formed from primary compounds [47,109,110]. Some of these metabolites are produced and stored during the normal growth of plants, whereas others accumulate only in response to stress conditions [111].
Due to their strong bioactivity, aromatic plants and their constituents may be applied as part of environmentally friendly management programs [64,112] to improve soil quality [113] and protect plants from pathogens and insects [114]. In a cropping system, aromatic plants and their bioactive metabolites can affect phytopathogens directly or indirectly by inducing systemic resistance and defense responses, repelling viral vectors, or inhibiting the growth of phytopathogens by changing the structure of soil microbial community [115,116,117,118,119]. Therefore, introduction of these noncrop biocontrol plants can affect crop productivity [120], with the postextraction residues and other plant debris being further sources of bioactive components that can be recycled as natural pesticides, organic amendments or transformed into value-added products providing nutrients, organic matter, and beneficial microorganisms [64,121,122]. For these reasons, the following sections provide background information on the applicability of aromatic and medicinal plants, botanicals, postextraction residues, and composts derived from aromatic plant waste to control the spread of major soil-borne pathogens.
4.1. Use of Pure Extracts as Biopesticides
Microorganisms are considered to be the most important biopesticides in the fight against plant diseases, whereas botanicals still represent a relatively small part of the market. Prior to World War II, only four major components were commonly used, i.e., rotenone and rotenoids, pyrethrum and pyrethrins, nicotine and alkaloids, and some vegetable oils [123], while other allelochemicals, such as derris and citronella oil, were applied as insecticides [90]. Although their use declined following the commercialization of synthetic agrochemicals due to their instability (pyrethrum) and toxicity to nontarget species (rotenone and nicotine) [123,124], the multitudinal backlash against synthetic compounds eventually led to the prosperity of biopesticides of plant origin [125].
The group of botanicals includes various plant volatiles, extracts, and natural oils that are characterized by improved biocompatibility, little or no cross-resistance to commercial chemical agents, and structural diversity [126,127]. Generally, these biologically active substances are extracted or distilled from fresh or dried plant materials with classical methods using water, alcohol, and other solvents [127]. To decrease the environmental impact, green processes requiring no or low amounts of solvents and less energy and time have also been developed including subcritical water extraction, pressurized ultrasound and microwave extraction, and supercritical fluid extraction [128,129,130].
4.1.1. Essential Oils
Essential oils (EOs), biosynthesized, stored, and secreted in various parts of aromatic herbs, e.g., buds, bark, leaves, zest, twigs, wood, stems, seeds, fruits, rhizome, roots, and flowers [128,131,132,133], are complex mixtures of terpenoids and phenolic compounds that play a critical role in plant defense mechanisms against abiotic and biotic stress factors [128,132,134]. These volatile organic compounds can diffuse through water- and gas-filled pores and serve as food sources, infochemicals, chemo-attractants, or antimicrobials [135]. Monoterpenes, as frequent constituents of EOs, are involved in chemical and ecological interactions, and their production may be induced by the attack of pathogens and herbivores [136,137]. Due to their bioactive potential, EOs and their constituents can be utilized as components of ecological products with nematicidal, antimicrobial, antiviral, insecticidal, repellent, antifeedant, and molluscicidal activity [23,118,138]. They can also support the growth of other soil microbes indirectly by killing organisms and providing easily decomposable carbon and energy sources [139]. This was confirmed by [140], who found that Mentha × piperita EO had an instant antimicrobial effect on soil; however, a fraction of soil microbes survived the treatment and used the killed microorganisms as fresh organic carbon. As a result, an increased microbial biomass and respiration rate were recorded.
As for the efficiency of active substances derived from aromatic and medicinal plants, EOs tend to possess stronger antimicrobial properties than other extracts [51]. Although several studies investigated the biocidal effect of EOs and their constituents on phytopathogens [46,141,142], the mechanism of their action is complex and not fully understood [143,144]. Volatile oils containing oxygenated monoterpenes, primarily phenols and aldehydes, tend to be more active against pathogens [145,146]. This was confirmed by Wogiatzi et al. [147], who found that the thymol-rich EO of Origanum vulgare collected from the mountain Olympus in Greece had the strongest antifungal effect on Verticillium dahliae, Pythium spp., and Sclerotinia sclerotiorum. Conversely, the hydrocarbonic fraction of Salvia pomifera L. ssp. calycina (Sm.) Hayek was less effective against Sclerotinia sclerotiorum and Rhizoctonia solani than was the oxygenated fraction [148].
The application of EOs can provoke chemical, physical, and biochemical changes in the cell [149] including disruption of the permeability barrier and proton pump function, damage of cytoplasmic membrane, increased permeability and leakage of cell contents, decreased ATP synthesis and augmented ATP hydrolysis, cytoplasm coagulation, reduced membrane potential, and inhibition of the production of toxic microbial metabolites [133,150]. As regards fungi, EOs can inhibit ergosterol biosynthesis leading to the depletion of sterol content in the cell membrane, disruption of cell membrane integrity and permeability, loss of ions, and, eventually, inhibition of fungal growth [151]. The study of Upadhyay et al. [151] has shown that a decreased level of methylglyoxal may also be observed that was reported to be in correlation with aflatoxin B1 production in Aspergillus flavus. In addition, Vokou et al. [139] claimed that volatile oils extracted from Satureja thymbra could reduce the spore germination and mycelial growth of Penicillium citrinum and Mucor hiemalis, respectively. Another study conducted by Sempere-Ferre et al. [152] showed that EO constituents, either alone or in combination, inhibited the mycelial growth of Botryotinia fuckeliana and Rhizoctonia solani, although eugenol exerted a fungistatic effect only. Similarly, testing the antifungal efficacy of various EO constituents, Marei et al. [153] found that 1,8-cineole, (R)-camphor, (R)-carvone, camphene, cuminaldehyde, (R)-linalool, geraniol, (1R,2S,5R)-menthol, (S)-fenchone, myrcene, thymol, and (S)-limonene were promising substances against Fusarium oxysporum, Penicillium digitatum, Aspergillus niger, and Rhizoctonia solani. Thymol and (S)-limonene possessed strong inhibitory effect on cellulase and pectin methyl esterase activities. Lee et al. [154] stated that Origanum vulgare EO had a broad antifungal spectrum against both postharvest (Botrytis cinerea, Colletotrichum gloeosporoides) and soil-borne pathogenic fungi (Fusarium oxysporum, Rhizoctonia solani, and Pythium ultimum). Bi et al. [155] reported that a 21-day-long soil treatment with EOs from palmarosa (Cymbopogon martini), oregano (Origanum syriacum), and red thyme (Thymus vulgaris) at concentrations of ≥0.1 μg/mL decreased the population density of Phytophthora capsici below the limit of detection.
As nematicides, EOs can reduce gall formation, hinder the hatching of nematode eggs, immobilize juveniles, and, as a result, completely suppress nematode infestation [156]. This is consistent with the findings of Ntalli et al. [65], who reported the EOs of Origanum vulgare, Origanum dictammus, Mentha pulegium, and Melissa officinalis to be effective against Meloidogyne incognita by irreversibly paralyzing the larvae (Table 1). In fact, the oxygenated EO constituents were more active than hydrocarbons, including β-caryophyllene, p-cymene, and limonene.
Overall, the studies presented thus far provide evidence that the biological activity of EOs is usually strongly related to their chemical composition; the synergistic effect of different bioactive components [65,148]; specific seasonal, geographic, and climatic conditions [64]; genetic factors; and harvest and postharvest processes [157].
4.1.2. Phenolic Compounds
Aromatic plants are rich sources of phenolic compounds (e.g., phenols, coumarins, tannins, flavonoids, phenolic acids) derived from the phenylpropanoid metabolism and shikimate pathway [158,159]. So far, around 8000 phenolics with highly diverse structures have been reported in the literature [160,161]. They contain one or more aromatic rings (C6) bearing at least one hydroxyl group [162,163]. These compounds are toxic to fungi, bacteria, insects, nematodes, and weeds [164], but, unlike EOs, they are water-soluble because they mostly exist in glycosidic form in plants [165].
Although polyphenols were isolated from various plant parts [166], they are found at higher levels in the outer layers of tissues [167] as the production of these metabolites can be induced by both biotic and abiotic factors such as reactive oxygen and nitrogen species, insufficient amounts of nutrients, low temperatures, UV light, predators, parasites, pathogens, wounding, and other stress conditions [163,168,169]. Nevertheless, the antimicrobial potential of polyphenols has not been fully deciphered [170] due to the complexity of their natural mixtures in plants [171] and their inconsistent activity [172]. Among them, flavonoids are associated with the stronger biocidal effects, whereas nonflavonoid compounds usually show a weak antagonistic behavior against pathogens [173]. Their activity, however, depends not only on their purity, structure, and concentration but also on the tested microbial strains, solvents, and experimental conditions [172].
Natural phenolics combine seemingly antagonistic effects, including an antioxidant activity, with variable degrees of cytotoxicity [174]. The mode of action of phenolics involves modification of cell membrane permeability and changes in intracellular functions, such as [170] inhibition of efflux pump and enzymatic (i.e., DNA gyrase, beta-ketoacyl acyl carrier protein synthase II and III, FabG, FabZ, FabI, d-alanine–d-alanine ligase, urease, sortase A, dihydrofolate reductase, etc.) activities, leakage of intracellular constituents, disturbance of cell wall biosynthesis, and inhibition of bacterial biofilm formation [175,176]. Important phenolics can repress or induce the expression of genes playing important roles in the pathogenesis of soft-rot-causing pathogenic bacteria or induce genes encoding efflux pumps in soft-rot enterobacteria [174]. As Summers and Felton [177] stated, phenolics could result in reduced protein digestibility, impaired enzymatic functions, and reduced bioavailability of amino acids by forming complexes with proteins. In addition, Simmonds [178] mentioned that the ingestion of phenolic compounds can reduce the nutritive value of the food consumed by nonadapted insects. This effect may be due to the induction of oxidative stress to the digestive system [177]. Widmer and Laurent [179] reported that lavender (Lavandula angustifolia), lavender hybrid (Lavandula angustifolia × Lavandula spicata), and rosemary (Rosmarinus officinalis) leaf extracts reduced the germination of Phytophthora capsici, Phytophthora megakarya, and Phytophthora palmivora zoospores. The aqueous extract of Acacia saligna H. L. Wendl. containing benzoic acid, caffeine, o-coumaric acid, naringenin, quercetin, and kaempferol effectively inhibited the mycelial growth of Fusarium culmorum, Penicillium chrysogenum, and Rhizoctonia solani, especially at high concentrations (Table 2). At the same time, the minimal inhibitory concentration of the extract was 200, 300, 300, and 100 µg/mL against Agrobacterium tumefaciens, Enterobacter cloacae, Erwinia amylovora, and Pectobacterium carotovorum subsp. carotovorum, respectively [180]. The filter-sterilized water extracts of Origanum vulgare L., Melissa officinalis L., and Salvia officinalis L. shoots (0.5 and/or 2.0% w/v) suppressed the propagation of Fusarium oxysporum f. sp. asparagi [181]. Similar findings were reported by Ahmad and Matsubara [182], who demonstrated that the aqueous extract of Thymus vulgaris L. (0.5% and 2.0%) had the maximum suppression effect on Fusarium oxysporum f. sp. cyclaminis. Caffeic acid and rosmarinic acid, as the main constituents, also exhibited a stronger antifungal activity. To induce the defense responses of soybean and sorghum, Colpas et al. [112] used the aqueous extracts of Ocimum gratissimum leaves, which caused an instant increase in phytoalexin production. López et al. [183] demonstrated that the dichlormethanolic extracts of Melissa officinalis L. subsp. officinalis, Mentha longifolia (L.) Hudson, Origanum vulgare L. subsp. virens Bonnier & Layens, and Salvia pratensis L., as well as the ethyl acetate extract of Mentha × piperita L., Salvia pratensis L., and Thymus praecox Opiz subsp. Polytrichus inhibited the growth of Rhizopus stolonifer over 20% under in vitro conditions. The nematicidal activity of aromatic plant extracts was demonstrated by Ntalli et al. [64]: Origanum vulgare L. and Thymus citriodorus (Schreb) extracts were effectively utilized against Meloidogyne javanica (Treub) and Meloidogyne incognita (Kofoid and White). However, elevated concentrations of the active constituents may have an inhibitory effect on physiological processes of crop plants by increasing electrolyte leakage from the cell membrane of seedlings and negatively affecting seed germination, chlorophyll synthesis, and the functioning of photosystem II [184].
4.1.3. Applicability of Pure Metabolites in Plant Protection
Plant-derived biopesticides are inexpensive and readily available materials, which do not contaminate the environment due to their biodegradable nature [47,127]. Despite the extensive body of scientific literature and the tremendous amount of experience that support the bioactivity of botanicals on plant pathogens, only a few botanical pesticides are available and utilized in agriculture due to the lack of field verification [127,185]. In Hungary, for instance, the following plant-based pesticides and plant conditioners have been approved: Green NanoherbTM and Biomit® with a blend of EOs, DU-OLTM with lavender and lemon oil, ORGANICTM with orange oil, and HerbalTM containing herbal extracts [186,187].
Although several plant-based protectants, e.g., spearmint oil and clove oil, have even been included in the pesticide database of the European Union and approved by the European Commission [149,188], the commercialization of botanical extracts has been limited by regulatory policies, costs, and prolonged procedures [47]. These bioactive substances can hinder the growth of soil-borne pathogens with a relatively mild effect on the nontarget species [35]. However, they are prone to decompose rapidly, whereas moisture, air, or sunlight may affect their stability, efficiency, and shelf-life. In addition, there are further drawbacks that can complicate the applicability of botanical extracts in the field, such as their nonselective nature, varying performance, and unavailability during certain parts of the growing season, as well as the absence of legislation; established residue tolerance; scientific validation; and sufficient data on stability, quality, and effect on human health [127]. The applicability of volatiles or other extracts may be also compromised by their dose-dependent properties [53,184].
Overall, plant-based bioactive substances may be integrated into common practices to reduce the use of conventional chemical treatments [127]. Formulation with emulsifying agents, polymers, surfactants, stabilizers, defoamers, solvents, and other substances can be used to ensure their stability, improve their efficiency, and control the release of biologically active components under field conditions [114].
4.2. Aromatic Plants as Part of Agricultural Practices: Cultivating Aromatic Plants and Recycling the Waste as Green Manure and Compost
The aerial parts and roots of aromatic plants and their waste can affect the composition of microbial populations in soil by releasing complex mixtures of chemicals (Figure 2). These secondary metabolites, excreted directly into the soil from the living and decomposing tissues [109,189], may inhibit the growth of phytopathogens [190].
Over the last decades, various forms of these herbs including cultivated plants, green manure, compost, and wastes from EO extraction (plant residues and hydrosol) have been tested as potential non-chemical treatments against soil-borne plant pathogens. In the next subsections, the potential effects of aromatic plants, their postextraction wastes, and compost on phytopathogens will be discussed.
4.2.1. Diversifying Cropping Systems with Aromatic and Medicinal Plants
As continuous cropping systems have been constrained by various biotic and abiotic factors (i.e., reduced soil quality, autotoxicity of the crop, shifts in microbial communities, reduced microbial diversity and abundance, prosperity of plant pathogens, etc.) [19], agricultural diversity has become a critical part of sustainable agroecosystems [191]. Diversification requires the combination of environmental and economic sustainability through increasing crop species diversity (intercropping, crop rotation) or planting noncrops (cover-cropping) [19,191,192,193]. These strategies may reduce production costs and improve resource use and soil quality [192], while the highly diverse plant communities thus created are less affected by phytopathogens [193]. However, the reduced activity of plant pathogens in such communities depends both on host density and the effects of neighboring species on disease-causing organisms [43].
Under optimal conditions, the phytomanagement of soils through the cultivation of aromatic and medicinal plant species can be a win-win approach that provides additional profits and marketable products [194,195,196]. As a vegetation cover, herbs can increase the fertility and the organic matter content of soils resulting in improved properties (e.g., water retention, soil porosity, permeability, and bulk density) [105].
It is also believed that the microbiota around medicinal and aromatic plants is highly plant-specific [196]. The rhizospheric zone in the vicinity of the plant roots is an attractive environment for a large number of organisms because plants may release up to 20% of the photosynthetically fixed carbon through their roots [119]. In addition to the typical root exudates, such as sugars, organic acids, amino acids, and sterols, aromatic crops can release novel chemicals in the soil that may have a long-lasting effect on the processes arbitrated by the surrounding microorganisms. Although the production of plant secondary metabolites is relatively low [110,116,197], volatiles can form a stable concentration gradient in the soil due to the lack of turbulence [109], and the build-up of pathogenic microorganisms and other predators can thus be inhibited [191]. However, several rhizospheric microorganisms have the potential to adapt and conduct endophytic or epiphytic life. These microbes and the host plant will influence each other through biological and physicochemical interactions [198]. For instance, the microorganisms associated with the roots of aromatic plants take part in plant growth and nutrition, regulate disease interactions [196], and may induce the production of certain bioactive compounds [117].
Over the past decades, several studies have confirmed the beneficial effects of aromatic herbs on soil microbial composition. Karthikeyan et al. [199] showed that the bacterial, fungal, and actinomycete populations in the rhizosphere of Ocimum sanctum L. were 2.3 × 107, 1.9 × 105, and 1.2 × 106 CFU/g, respectively. These values were higher than the corresponding ones observed in the rhizosphere of Coleus forskholii Briq. Similar results were reported for diazotrophic bacterial populations (i.e., Azospirillum spp., Azotobacter spp., and Pseudomonas spp.). Examining soil microbial populations, Adamović et al. [200] observed an increase in the total number of microorganisms, cellulolytic microorganisms, azotobacters, fungi, and free nitrogen-fixing microorganisms when mint (Mentha × piperita L.) was planted. The cultivation of basil (Ocimum basilicum L.) had the same effect on the total numbers of actinomycetes. Zhang et al. [113] showed that intercropping pear trees with basil or summer savory (Satureja hortensis L.) could increase the organic matter content of soil and the activities of enzymes including invertase, urease, and catalase due to the influence of root exudates on soil microbial diversity. Meanwhile, soil organic matter was negatively correlated with pathotrophic groups of fungi, and, thus, intercropping significantly reduced the relative abundance of pathotrophic and saprotrophic groups. Bais et al. [111] reported that the fungal cell wall elicitors from Phytophthora drechsleri and Phytophthora cinnamomi improved root growth of Ocimum basilicum L. and facilitated rosmarinic acid production in hairy root cultures. Former studies have demonstrated that both EOs and aromatic water may possess antiphytoviral activity [80], but the physicochemical properties of soil and root exudates can also influence the migration and location of these organisms [74]. Due to their phytoremediation potential, several aromatic and medicinal plants are also used for the remediation of heavily contaminated soil by accumulating and removing organic and inorganic contaminants [201].
It should be noted that consecutive monocultures of herbs may severely alter the bacterial and fungal populations in soil [202]. According to the research conducted by Tang et al. [19], the continuous cropping of Salvia miltiorrhiza Bunge affected the microbial composition (e.g., fungal and actinomycete communities) of soil both in terms of structure diversity and abundance. Overall, the productivity of these plants tends to decrease over time because of the accumulation of their natural enemies [43] and the relatively low diversity of functional microbes [19].
Crop rotation is one of the best options to keep the levels of soil-borne pathogens and weeds low, but, from an economical perspective, it is not always acceptable [44,203]. Cover cropping and intercropping are used more and more frequently in conventional habitat management systems [138]. Aromatic herbs are excellent intercropping options for different cultures due to their characteristic traits and resistance to adverse environmental conditions [204]. Verma et al. [192] found that intercropping Pelargonium graveolens L. with companion crops may improve the postharvest soil total Kjeldahl nitrogen; organic carbon; carbon-to-nitrogen ratio; and available N, P, and K contents. The cultivation of wheat, oat, and barley also enhanced the biomass-specific respiration of soil that might be in association with higher microbial activity. Khan et al. [194] reported that cropping treatments with Ocimum basilicum L. cv. CIM-Saumya and Mentha arvensis L. cv. Kosi along with crop residue retention showed reduced CO2-C emission and increased soil organic carbon content compared to fallow soil. Chand et al. [205] showed that the cultivation of Pelargonium graveolens, Rosmarinus officinalis, and Mentha piperita had a conservation function, and these herbs reduced soil erosion and slowed down the runoff. However, the relative soil and water conservation efficiency of Thymus vulgaris was almost zero due to its poor canopy coverage. The authors also suggested that geranium and rosemary were good candidates for vegetative barriers, intercrops, and cover crops.
The plant material of herbs can be left on the top of the soil as mulch or incorporated into green manure [206]. In this form, they can interfere with phytopathogens directly through the release of toxic compounds or indirectly [207] by favoring the natural enemies of soil-borne pathogens through habitat manipulation [35]. Furthermore, the supplementation of plant growth-promoting microorganisms associated with medicinal plants can improve plant health by increasing their immunity to phytopathogens [208], improving stress tolerance, and influencing nutrient availability and uptake, as well as the production of growth-promoting metabolites and hormones, such as cytokinins and auxins [209]. Castronovo et al. [210] reported that microorganisms isolated from the bulk soil of Origanum vulgare L. exhibited antibacterial activity against various human pathogens. The Agrobacterium, Agromyces, Bacillus, and Chryseobacterium isolates inhibited the growth of several Bacillus strains of environmental origin. Tiwari et al. [211] isolated potential plant growth-promoting bacterial agents (Bacillus and Pseudomonas spp.) from the root vicinity of medicinal aromatic plants such as Ocimum spp. Under greenhouse conditions, the isolated bioinoculants alone or in combination with Trichoderma harzianum reduced the reproduction factor of Meloidogyne incognita by 46.4–72.3% in sterile and natural soils. Chowdhary and Kaushik [212] reported that approximately 12% of endophytic fungal isolates from Mentha × piperita plants exhibited antifungal activity against three or more of the tested phytopathogens.
In addition to the aforementioned beneficial effects on soil microbial composition, aromatic plant-based cropping systems can provide significantly increased profits and crop production rates [213] because herbs (e.g., Ocimum basilicum L.) are less prone to compete with cultivated crops for nutrients [214,215]. Carvalho et al. [216] studied how intercropping with aromatic plants, such as Ruta graveolens, can affect tomato yield. This cultivation technique was found to increase the total yield of tomato fruits by 26%. In a similar companion planting experiment, Ahmad et al. [214] observed that certain Lamiaceae species (e.g., Ocimum basilicum L., Mentha piperita L., Hyssopus officinalis L.) promoted the growth of tomato plants and influenced the production and accumulation of certain metabolites in the dominant crop. Peppermint and hyssop enhanced shikimic acid and apigenin contents in all parts of tomato plants providing a product with improved quality and yield. Likewise, cropping with basil resulted in higher free amino acid contents with the leaves being rich in alanine, phenylalanine, iso-leucine, lysine, valine, and γ-aminobutyric acid (GABA), whereas the stems contained elevated levels of valine, serine, alanine, proline, GABA, and glutamine compared to the control treatment. In addition, the 1:1 tomato–aromatic plant companion setups boosted the growth of tomato plants without the accumulation of competition pressure. Under certain circumstances, however, temporal phytotoxic effects could be experienced due to the incorporation of aromatic herb biomass. Nevertheless, other techniques (e.g., relay-planting) are available that can optimize productivity [217].
4.2.2. Recycling the By-Products of Aromatic Plant-Utilizing Sectors
The demand for aromatic and medicinal plant-based products has increased continuously, at the rate of 15–25%, for the last few years [218]. As the ratio between the processed EO and the utilized plant material is low [219], large quantities of solid waste are generated and remain unutilized [220]. From a sustainability perspective, the aromatic biomass cannot be considered as waste [221], and it may be recycled as animal feed, biosorbents, biogas, biofuel, chelating agents, biopesticides, or soil amendments [121,219].
Just as with the fresh plant material, the distilled solid biomass can contain volatile components and phenolic compounds, albeit in different ratios [219,222]. According to Kadoglidou et al. [33], the application of Mentha spicata L. and Origanum vulgare L. ssp. hirtum plant material as soil amendments (4%) decreased the average degree of infection (Fusarium oxysporun f. sp. lycopersici and Verticillium dahliae) of tomatoes. In addition to plant disease reduction, the growth and physiology indices as well as soluble solids contents and fruit yield were also affected. Moreover, the amendment of Mentha spicata L. tissue at different concentrations may have a positive impact on tomato seedling emergence and dry weight, the size of the most robust leaf, shoot length, physiological parameters (e.g., stomatal conductance, photosynthetic rate, and yield), and the size of fungal and bacterial populations [223]. Klein et al. [224] used herbal biomass in combination with soil solarization to control the spread of Fusarium oxysporum f. sp. radicis-lycopersici and Meloidogyne javanica under farm conditions. The results showed that solarization with rosemary (Rosmarinus officinalis L.), sage (Salvia officinalis L.), tarragon (Artemisia dracunculus L.), and thyme (Thymus vulgaris L.) residues was highly effective against the tested fungi reducing their viability by 99–100%. As for the root-knot nematode, galling was reduced to zero in tomato roots in the solarized soil amended with sage and thyme residues. Despite the positive effect of untreated herbal residues on soil productivity and disease severity, their direct use may be compromised due to their unknown composition [157]. Furthermore, the introduction of novel plant metabolites may also affect indigenous microbial communities [116], critically the decomposition process [225], and endanger crop growth and yield [203]. As an example of the negative impact, Gravanis et al. [226] reported that the incorporation of dried Origanum vulgare biomass significantly decreased the number of bacterial colonies in soil samples. Similar results were reported by Chouliaras et al. [227]. In contrast, Ainalidou et al. [228] showed that soil enrichment with the green parts of Mentha spicata, Mentha piperita, and Rosmarinus officinalis increased the total microbial biomass. However, the use of rosemary had a negative effect on the growth indices of tomato seedlings, including weight, shoot length, and root length of fresh seedlings, as also reported by Argyropoulou et al. [229].
The meaningful utilization of aromatic biomass is still a great challenge. Composting and vermitechnology are commonly applied to treat these agricultural wastes prior to use. The mature and stable end product is a suitable soil amendment that enhances important soil processes and features improving the diversity and activity of microbial communities in soil [223,230]. Moreover, composts provide a new environmental source of microbes with antimicrobial activity [231]. Although their efficiency is variable, the microorganisms involved in the decomposition of organic matter can suppress the growth of soil-borne pathogenic microbes through antagonistic interactions, inducing systemic resistance in the host plant or forming humic molecules and biostimulants [232]. Under glasshouse conditions, the nematicide activity of vermicomposts containing Artemisia annua, Chrysanthemum cinerariaefolium, Plantago ovata, Mentha arvensis, Pelargonium graveolens, and Tagetes minuta was examined by Pandey and Kalra [233]. The maximum reduction in root galling was recorded in tomato plants treated with menthol mint vermicompost [Root-Knot Index (RKI): 1.33] followed by marigold (RKI: 1.66), isabgol (RKI: 1.66), and qinghao (RKI: 1.81). Compared to one of the most common chemical nematicides (carbofuran), increased fresh and dry root and shoot weights and fruit yields were achieved with certain herb-based vermicompost treatments. Singh et al. [234] prepared vermicompost from distillation waste of aromatic oil crops (e.g., Cymbopogon flexuosus and Cymbopogon winterianus) with Eisenia fetida to control root-rot of Coleus forskohlii caused by Fusarium chlamydosporum and Ralstonia solanacearum. Zhou et al. [235] co-composted food waste and sawdust with Chinese medicinal herb residues (1:1:1). The acetone extract of the end products showed antagonistic activity against Fusarium oxysporum and Alternaria solani, but the antimicrobial properties of the mature compost were as strong as those of the Chinese herbal residues only. It was concluded that the growth inhibition observed was partly due to the natural microbiota of the compost. In eleven composts, Zaccardelli et al. [122] found 104 spore-forming bacterial isolates that exerted antagonistic effects on Sclerotinia minor and Rhizoctonia solani. Among the seven most promising isolates, the in vivo antipathogenic activity of two Bacillus subtilis strains and a Bacillus amyloliquefaciens isolate was confirmed against Sclerotinia minor on Diplotaxis tenuifolia L.
In addition to the solid by-products, the hydrolates remaining after the extraction process may also possess biocidal properties [236] as a result of leaching of certain bioactive compounds (i.e., trace amounts of EOs and other water-soluble components) during hydrodistillation [219,237]. The hydrosol extract of Thymus capitatus L. rich in carvacrol effectively inhibited the growth of important fungal pathogens of Citrus sinensis L. including Aspergillus niger, Aspergillus oryzae, and Fusarium solani [238]. Gaspar-Pintiliescu et al. [239] reported antibacterial activity for the aromatic water of Rosmarinus officinalis. Under in vitro conditions, strong nematicidal activity was associated with the use of hydrolates derived from Lavandula × intermedia Emeric ex Loisel. var. super and Lavandula luisieri (Rozeira) Rivas-Martínez [240]. A similar effect of Thymus citriodorus (Schreb) was demonstrated by Ntalli et al. [241] against Meloidogyne incognita and Meloidogyne javanica. Sainz et al. [242] reported a high mortality rate for second-stage juveniles of Meloidogyne javanica upon use of a hydrolate from Artemisia pedemontana subsp. assoana. However, hydrolate-based natural pesticides may cause acute toxicity at higher doses for non-target organisms, reduce the metabolism of the natural soil microbiota, and slightly influence the physiological diversity of microbial communities [243].
5. Conclusions
Modern agriculture still relies heavily on the application of synthetic chemicals, even though controversies have arisen regarding their safety. Due to the public’s concerns, the strict regulations, and the pursuit of agricultural sustainability, low-risk and environmentally compatible biocontrol alternatives have emerged as suitable substitutes for these agrochemicals. Botanicals, including EOs and phenolic compounds, have been used for decades as non-chemical treatments thanks to their biodegradable nature and high selectivity. Their market is continuously growing; however, they only make up a small percentage of the global market, and just a few of these aromatic plant-based biopesticides are commercially available because of their varying performance and the absence of field studies regarding their efficiency. The inclusion of aromatic and medicinal herbs into agricultural practices is a possible way to increase crop diversity. In addition, the use of their industrial by-products as organic fertilizers may suppress the growth of major soil-borne phytopathogens. The cultivation of aromatic herbs can also improve crop production rates and the chemical composition of other plants. If used as part of integrated pest management programs, they are capable of reducing the environmental impacts of chemical pesticides. In the last few decades, disposal of by-products (e.g., solid biomass, aromatic water, etc.) generated by the aromatic and medicinal plant-utilizing sector has become a common issue throughout the world. Since these wastes may be rich sources of bioactive compounds and beneficial microorganisms, their use as natural biopesticides or organic amendments has also gained popularity. Composting and vermicomposting technologies can offer a meaningful way to produce a mature and stable end product that can improve important soil processes and provide nutrients for the treated crops.
All in all, botanicals, and especially EOs, are the most studied and commonly used treatments against soil-borne plant pathogens. There is substantial evidence that several of them possess strong antifungal properties, thereby reducing mycelial growth and/or spore germination of various fungal taxa such as Fusarium, Aspergillus, Penicillium, Rhizoctonia solani, Verticillium dahliae, and Sclerotinia sclerotiorum. Moreover, they show pronounced nematicidal activities against phytoparasitic, root-knot nematodes (Meloidogyne spp.). Nevertheless, the application of these methods is far behind their true potential due to some shortcomings, e.g., lack of in vitro experiments, varying selectivity, diversity of bioactive compounds and complex mechanisms of action, rigorous approval procedures and registration processes, etc. Therefore, more insight is required to overcome these obstacles and improve the overall commercialization process of plant-based biocontrol agents.
Author Contributions
Conceptualization, B.G., A.S., E.L. and L.V.; methodology, B.G. and L.V.; formal analysis, L.V.; investigation, B.G. and A.S.; resources, E.L.; writing—original draft preparation, B.G. and A.S.; writing—review and editing, L.V.; supervision, E.L.; project administration, E.L.; funding acquisition, E.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the ÚNKP-22-4-II-SZE-24 New National Excellence Program of the Ministry for Culture and Innovation (Hungary) from the source of the National Research, Development, and Innovation Fund.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gomiero, T. Soil degradation, land scarcity and food security: Reviewing a complex challenge. Sustainability 2016, 8, 281. [Google Scholar] [CrossRef]
- Nambiar, K.K.M.; Gupta, A.P.; Fu, Q.L.; Li, S. Biophysical, chemical and socio-economic indicators for assessing agricultural sustainability in the Chinese coastal zone. Agric. Ecosyst. Environ. 2001, 87, 209–214. [Google Scholar] [CrossRef]
- Trivedi, P.; Delgado-Baquerizo, M.; Anderson, I.C.; Singh, B.K. Response of soil properties and microbial communities to agriculture: Implications for primary productivity and soil health indicators. Front. Plant Sci. 2016, 7, 990. [Google Scholar] [CrossRef] [PubMed]
- Turmel, M.S.; Speratti, A.; Baudron, F.; Verhulst, N.; Govaerts, B. Crop residue management and soil health: A systems analysis. Agric. Syst. 2015, 134, 6–16. [Google Scholar] [CrossRef]
- Congreves, K.A.; Hayes, A.; Verhallen, E.A.; Van Eerd, L.L. Long-term impact of tillage and crop rotation on soil health at four temperate agroecosystems. Soil Tillage Res. 2015, 152, 17–28. [Google Scholar] [CrossRef]
- Doran, J.W.; Zeiss, M.R. Soil health and sustainability: Managing the biotic component of soil quality. Appl. Soil Ecol. 2000, 15, 3–11. [Google Scholar] [CrossRef]
- De Corato, U. Disease-suppressive compost enhances natural soil suppressiveness against soil-borne plant pathogens: A critical review. Rhizosphere 2020, 13, 100192. [Google Scholar] [CrossRef]
- van Bruggen, A.H.C.; Semenov, A.M. In search of biological indicators for soil health and disease suppression. Appl. Soil Ecol. 2000, 15, 13–24. [Google Scholar] [CrossRef]
- De Corato, U. Soil microbiota manipulation and its role in suppressing soil-borne plant pathogens in organic farming systems under the light of microbiome-assisted strategies. Chem. Biol. Technol. Agric. 2020, 7, 17. [Google Scholar] [CrossRef]
- Hannula, S.E.; Ma, H.; Pérez-Jaramillo, J.E.; Pineda, A.; Bezemer, T.M. Structure and ecological function of the soil microbiome affecting plant–soil feedbacks in the presence of a soil-borne pathogen. Environ. Microbiol. 2020, 22, 660–676. [Google Scholar] [CrossRef] [Green Version]
- Mendes, L.W.; Mendes, R.; Raaijmakers, J.M.; Tsai, S.M. Breeding for soil-borne pathogen resistance impacts active rhizosphere microbiome of common bean. ISME J. 2018, 12, 3038–3042. [Google Scholar] [CrossRef]
- Wolfgang, A.; Taffner, J.; Guimarães, R.A.; Coyne, D.; Berg, G. Novel strategies for soil-borne diseases: Exploiting the microbiome and volatile-based mechanisms toward controlling Meloidogyne-based disease complexes. Front. Microbiol. 2019, 10, 1296. [Google Scholar] [CrossRef] [PubMed]
- Haas, D.; Défago, G. Biological control of soil-borne pathogens by fluorescent pseudomonads. Nat. Rev. Microbiol. 2005, 3, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Lambraño, R.E.; González-Moreno, P.; Sánchez-Agudo, J.Á. Environmental factors associated with the spatial distribution of invasive plant pathogens in the Iberian Peninsula: The case of Phytophthora cinnamomi Rands. For. Ecol. Manag. 2018, 419–420, 101–109. [Google Scholar] [CrossRef]
- Katan, J. Diseases caused by soilborne pathogens: Biology, management and challenges. J. Plant Pathol. 2017, 99, 305–315. [Google Scholar] [CrossRef]
- Mehta, C.M.; Palni, U.; Franke-Whittle, I.H.; Sharma, A.K. Compost: Its role, mechanism and impact on reducing soil-borne plant diseases. Waste Manag. 2014, 34, 607–622. [Google Scholar] [CrossRef]
- Li, X.G.; Zhang, T.L.; Wang, X.X.; Hua, K.; Zhao, L.; Han, Z.M. The composition of root exudates from two different resistant peanut cultivars and their effects on the growth of soil-borne pathogen. Int. J. Biol. Sci. 2013, 9, 164–173. [Google Scholar] [CrossRef]
- van Elsas, J.D.; Garbeva, P.; Salles, J. Effects of agronomical measures on the microbial diversity of soils as related to the suppression of soil-borne plant pathogens. Biodegradation 2002, 13, 29–40. [Google Scholar] [CrossRef]
- Tang, J.; Xue, Z.Q.; Daroch, M.; Ma, J. Impact of continuous Salvia miltiorrhiza cropping on rhizosphere actinomycetes and fungi communities. Ann. Microbiol. 2015, 65, 1267–1275. [Google Scholar] [CrossRef]
- Bailey, K.L.; Lazarovits, G. Suppressing soil-borne diseases with residue management and organic amendments. Soil Tillage Res. 2003, 72, 169–180. [Google Scholar] [CrossRef]
- Du, F.Y.; Ju, G.L.; Xiao, L.; Zhou, Y.M.; Wu, X. Sesquiterpenes and cyclodepsipeptides from marine-derived fungus Trichoderma longibrachiatum and their antagonistic activities against soil-borne pathogens. Mar. Drugs 2020, 18, 165. [Google Scholar] [CrossRef] [PubMed]
- Panth, M.; Hassler, S.C.; Baysal-Gurel, F. Methods for management of soilborne diseases in crop production. Agriculture 2020, 10, 16. [Google Scholar] [CrossRef]
- Barua, A.; McDonald-Howard, K.L.; Mc Donnell, R.J.; Rae, R.; Williams, C.D. Toxicity of essential oils to slug parasitic and entomopathogenic nematodes. J. Pest Sci. 2020, 93, 1411–1419. [Google Scholar] [CrossRef]
- Cheng, H.Y.; Zhang, D.Q.; Ren, L.R.; Song, Z.X.; Li, Q.J.; Wu, J.J.; Fang, W.S.; Huang, B.; Yan, D.D.; Li, Y.; et al. Bio-activation of soil with beneficial microbes after soil fumigation reduces soil-borne pathogens and increases tomato yield. Environ. Pollut. 2021, 283, 117160. [Google Scholar] [CrossRef] [PubMed]
- Stuart, A.K.C.; Stuart, R.M.; Pimentel, I.C. Effect of agrochemicals on endophytic fungi community associated with crops of organic and conventional soybean (Glycine max L. Merril). Agric. Nat. Resour. 2018, 52, 388–392. [Google Scholar] [CrossRef]
- Ashraf, S.; Zuhaib, M. Fungal biodiversity: A potential tool in plant disease management. In Management of Microbial Resources in the Environment; Malik, A., Grohmann, E., Alves, M., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 69–90. [Google Scholar] [CrossRef]
- Verchot-Lubicz, J. Soilborne viruses: Advances in virus movement, virus induced gene silencing, and engineered resistance. Physiol. Mol. Plant Pathol. 2003, 62, 55–63. [Google Scholar] [CrossRef]
- Colla, P.; Gilardi, G.; Gullino, M.L. A review and critical analysis of the European situation of soilborne disease management in the vegetable sector. Phytoparasitica 2012, 40, 515–523. [Google Scholar] [CrossRef]
- Lougraimzi, H.; El Iraqui, S.; Bouaichi, A.; Gouit, S.; El Hassan, A.; Fadli, M. Insecticidal effect of essential oil and powder of Mentha pulegium L. leaves against Sitophilus oryzae (Linnaeus, 1763) and Tribolium castaneum (Herbst, 1797) (Coleoptera: Curculionidae, Tenebrionidae), the main pests of stored wheat in Morocco. Pol. J. Entomol. 2018, 87, 263–278. [Google Scholar] [CrossRef]
- Thampi, A.; Bhai, R.S. Rhizosphere actinobacteria for combating Phytophthora capsici and Sclerotium rolfsii, the major soil borne pathogens of black pepper (Piper nigrum L.). Biol. Control 2017, 109, 1–13. [Google Scholar] [CrossRef]
- Farzana; Abid, M.; Hussain, F. Screening of ethnomedicinal plants for their antifungal and nematicidal activities against soil-borne phytopathogens. S. Afr. J. Bot. 2022, 147, 18–23. [Google Scholar] [CrossRef]
- Govindarajan, M.; Kadaikunnan, S.; Alharbi, N.S.; Benelli, G. Acute toxicity and repellent activity of the Origanum scabrum Boiss. & Heldr. (Lamiaceae) essential oil against four mosquito vectors of public health importance and its biosafety on non-target aquatic organisms. Environ. Sci. Pollut. Res. 2016, 23, 23228–23238. [Google Scholar] [CrossRef]
- Kadoglidou, K.; Chatzopoulou, P.; Maloupa, E.; Kalaitzidis, A.; Ghoghoberidze, S.; Katsantonis, D. Mentha and oregano soil amendment induces enhancement of tomato tolerance against soilborne diseases, yield and quality. Agronomy 2020, 10, 406. [Google Scholar] [CrossRef]
- Pretorius, J.C.; Zietsman, P.C.; Eksteen, D. Fungitoxic properties of selected South African plant species against plant pathogens of economic importance in agriculture. Ann. Appl. Biol. 2002, 141, 117–124. [Google Scholar] [CrossRef]
- Amoabeng, B.W.; Johnson, A.C.; Gurr, G.M. Natural enemy enhancement and botanical insecticide source: A review of dual use companion plants. Appl. Entomol. Zool. 2019, 54, 1–19. [Google Scholar] [CrossRef]
- Shafique, H.A.; Athar, M.; Ehteshamul-Haque, S.; Sultana, V. Management of soil-borne diseases of organic vegetables. J. Plant Prot. Res. 2016, 56, 221–230. [Google Scholar] [CrossRef]
- Tahat, M.M.; Alananbeh, K.M.; Othman, Y.A.; Leskovar, D.I. Soil health and sustainable agriculture. Sustainability 2020, 12, 4859. [Google Scholar] [CrossRef]
- Bonanomi, G.; De Filippis, F.; Cesarano, G.; La Storia, A.; Ercolini, D.; Scala, F. Organic farming induces changes in soil microbiota that affect agro-ecosystem functions. Soil Biol. Biochem. 2016, 103, 327–336. [Google Scholar] [CrossRef]
- Shokoohi, E.; Mashela, P.W.; Machado, R.A.R. Bacterial communities associated with Zeldia punctata, a bacterivorous soil-borne nematode. Int. Microbiol. 2022, 25, 207–216. [Google Scholar] [CrossRef]
- Bakker, P.A.H.M.; Berendsen, R.L.; Van Pelt, J.A.; Vismans, G.; Yu, K.; Li, E.; Van Bentum, S.; Poppeliers, S.W.M.; Gil, J.J.S.; Zhang, H.; et al. The soil-borne identity and microbiome-assisted agriculture: Looking back to the future. Mol. Plant 2020, 13, 1394–1401. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, J.; Liang, H.; Huang, J.; Chen, Z.; Nie, Y.J.; Wang, C.B.; Wang, Y.G. Manipulation of the rhizosphere microbial community through application of a new bio-organic fertilizer improves watermelon quality and health. PLoS ONE 2018, 13, e0192967. [Google Scholar] [CrossRef]
- Sarwar, M.; Aslam, M.; Sarwar, S.; Iftikhar, R. Different nematodes and plasmodiophorids as vectors of plant viruses. In Applied Plant Virology: Advances, Detection, and Antiviral Strategies; Awasthi, L.P., Ed.; Academic Press: London, UK, 2020; pp. 275–290. [Google Scholar] [CrossRef]
- van Ruijven, J.; Ampt, E.; Francioli, D.; Mommer, L. Do soil-borne fungal pathogens mediate plant diversity–productivity relationships? Evidence and future opportunities. J. Ecol. 2020, 108, 1810–1821. [Google Scholar] [CrossRef]
- Lucas, P. Diseases caused by soil-borne pathogens. In The Epidemiology of Plant Diseases; Cooke, B.M., Jones, D.G., Kaye, B., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 373–386. [Google Scholar] [CrossRef]
- Delgado-Baquerizo, M.; Guerra, C.A.; Cano-Díaz, C.; Egidi, E.; Wang, J.T.; Eisenhauer, N.; Singh, B.K.; Maestre, F.T. The proportion of soil-borne pathogens increases with warming at the global scale. Nat. Clim. Chang. 2020, 10, 550–554. [Google Scholar] [CrossRef]
- Moutassem, D.; Belabid, L.; Bellik, Y.; Ziouche, S.; Baali, F. Efficacy of essential oils of various aromatic plants in the biocontrol of Fusarium wilt and inducing systemic resistance in chickpea seedlings. Plant Prot. Sci. 2019, 55, 202–217. [Google Scholar] [CrossRef]
- Arora, H.; Sharma, A.; Poczai, P.; Sharma, S.; Haron, F.F.; Gafur, A.; Sayyed, R.Z. Plant-derived protectants in combating soil-borne fungal infections in tomato and chilli. J. Fungi 2022, 8, 213. [Google Scholar] [CrossRef]
- Baysal-Gurel, F.; Kabir, M.N.; Liyanapathiranage, P. Effect of organic inputs and solarization for the suppression of Rhizoctonia solani in woody ornamental plant production. Plants 2019, 8, 138. [Google Scholar] [CrossRef]
- Wang, M.; Ling, N.; Dong, X.; Zhu, Y.Y.; Shen, Q.R.; Guo, S.W. Thermographic visualization of leaf response in cucumber plants infected with the soil-borne pathogen Fusarium oxysporum f. sp. cucumerinum. Plant Physiol. Biochem. 2012, 61, 153–161. [Google Scholar] [CrossRef]
- Malandrakis, A.A.; Kavroulakis, N.; Chrysikopoulos, C.V. Use of copper, silver and zinc nanoparticles against foliar and soil-borne plant pathogens. Sci. Total Environ. 2019, 670, 292–299. [Google Scholar] [CrossRef]
- Erdoğan, O.; Çelik, A.; Zeybek, A. In vitro antifungal activity of mint, thyme, lavender extracts and essential oils on Verticillium dahliae Kleb. Fresenius Environ. Bull. 2016, 25, 4856–4862. [Google Scholar]
- Ayala-Doñas, A.; de Cara-García, M.; Talavera-Rubia, M.; Verdejo-Lucas, S. Management of soil-borne fungi and root-knot nematodes in cucurbits through breeding for resistance and grafting. Agronomy 2020, 10, 1641. [Google Scholar] [CrossRef]
- Farhang, V.; Amini, J.; Ebadollahi, A.; Sadeghi, G.R. Ocimum basilicum L. essential oil cultivated in Iran: Chemical composition and antifungal activity against three Phytophthora species. Arch. Phytopathol. Plant Prot. 2014, 47, 1696–1703. [Google Scholar] [CrossRef]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Lozano-Durán, R.; Macho, A.P. Insights into the root invasion by the plant pathogenic bacterium Ralstonia solanacearum. Plants 2020, 9, 516. [Google Scholar] [CrossRef] [PubMed]
- Djebaili, R.; Pellegrini, M.; Ercole, C.; Farda, B.; Kitouni, M.; Del Gallo, M. Biocontrol of soil-borne pathogens of Solanum lycopersicum L. and Daucus carota L. by plant growth-promoting actinomycetes: In vitro and in planta antagonistic activity. Pathogens 2021, 10, 1305. [Google Scholar] [CrossRef] [PubMed]
- de Haan, E.G.; Dekker-Nooren, T.C.E.M.; van den Bovenkamp, G.W.; Speksnijder, A.G.C.L.; van der Zouwen, P.S.; van der Wolf, J.M. Pectobacterium carotovorum subsp. carotovorum can cause potato blackleg in temperate climates. Eur. J. Plant Pathol. 2008, 122, 561–569. [Google Scholar] [CrossRef]
- Mwangi, M.; Bandyopadhyay, R.; Ragama, P.; Tushemereirwe, W.K. Assessment of banana planting practices and cultivar tolerance in relation to management of soilborne Xanthomonas campestris pv musacearum. Crop Prot. 2007, 26, 1203–1208. [Google Scholar] [CrossRef]
- Kantor, M.; Handoo, Z.; Kantor, C.; Carta, L. Top ten most important U.S.-regulated and emerging plant-parasitic nematodes. Horticulturae 2022, 8, 208. [Google Scholar] [CrossRef]
- Collange, B.; Navarrete, M.; Montfort, F.; Mateille, T.; Tavoillot, J.; Martiny, B.; Tchamitchian, M. Alternative cropping systems can have contrasting effects on various soil-borne diseases: Relevance of a systemic analysis in vegetable cropping systems. Crop Prot. 2014, 55, 7–15. [Google Scholar] [CrossRef]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-López, R.; Palomares-Rius, J.E.; Wesemael, W.M.L.; et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.T.; Wu, H.B.; Jiang, H.Y.; Zhang, L.; Zhang, Y.N.; Mao, L.G. Thiophenes from Echinops grijsii as a preliminary approach to control disease complex of root-knot nematodes and soil-borne fungi: Isolation, activities, and structure–nonphototoxic activity relationship analysis. J. Agric. Food Chem. 2019, 67, 6160–6168. [Google Scholar] [CrossRef]
- Nguyen, V.C.; Villate, L.; Gutierrez-Gutierrez, C.; Castillo, P.; Van Ghelder, C.; Plantard, O.; Esmenjaud, D. Phylogeography of the soil-borne vector nematode Xiphinema index highly suggests Eastern origin and dissemination with domesticated grapevine. Sci. Rep. 2019, 9, 7313. [Google Scholar] [CrossRef]
- Ntalli, N.G.; Ozalexandridou, E.X.; Kasiotis, K.M.; Samara, M.; Golfinopoulos, S.K. Nematicidal activity and phytochemistry of greek Lamiaceae species. Agronomy 2020, 10, 1119. [Google Scholar] [CrossRef]
- Ntalli, N.G.; Ferrari, F.; Giannakou, I.; Menkissoglu-Spiroudi, U. Phytochemistry and nematicidal activity of the essential oils from 8 Greek Lamiaceae aromatic plants and 13 terpene components. J. Agric. Food Chem. 2010, 58, 7856–7863. [Google Scholar] [CrossRef] [PubMed]
- Vieira, P.; Gleason, C. Plant-parasitic nematode effectors—Insights into their diversity and new tools for their identification. Curr. Opin. Plant Biol. 2019, 50, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Palomares-Rius, J.E.; Castillo, P.; Navas-Cortés, J.A.; Jiménez-Díaz, R.M.; Tena, M. A proteomic study of in-root interactions between chickpea pathogens: The root-knot nematode Meloidogyne artiellia and the soil-borne fungus Fusarium oxysporum f. sp. ciceris race 5. J. Proteomics 2011, 74, 2034–2051. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Daub, M.; Luigs, H.G.; Jansen, M.; Strauch, M.; Merhof, D. High-throughput phenotyping of nematode cysts. Front. Plant Sci. 2022, 13, 965254. [Google Scholar] [CrossRef]
- Khan, M.R. Nematode infestation, a potential threat to Indian forests. Indian Phytopathol. 2020, 73, 397–414. [Google Scholar] [CrossRef]
- Ankrom, K.E.; Franco, A.L.C.; Fonte, S.J.; Gherardi, L.A.; de Tomasel, C.M.; Andriuzzi, W.S.; Shaw, E.A.; Sala, O.E.; Wall, D.H. Ecto- and endoparasitic nematodes respond differently across sites to changes in precipitation. Oecologia 2020, 193, 761–771. [Google Scholar] [CrossRef]
- Sorribas, F.J.; Djian-Caporalino, C.; Mateille, T. Nematodes. In Integrated Pest and Disease Management in Greenhouse Crops; Gullino, M.L., Albajes, R., Nicot, P.C., Eds.; Springer: Cham, Switzerland, 2020; pp. 147–174. [Google Scholar] [CrossRef]
- Abad, P.; Gouzy, J.; Aury, J.M.; Castagnone-Sereno, P.; Danchin, E.G.J.; Deleury, E.; Perfus-Barbeoch, L.; Anthouard, V.; Artiguenave, F.; Blok, V.C.; et al. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909–915. [Google Scholar] [CrossRef]
- Dong, L.Q.; Zhang, K.Q. Microbial control of plant-parasitic nematodes: A five-party interaction. Plant Soil 2006, 288, 31–45. [Google Scholar] [CrossRef]
- Wu, H.M.; Yan, W.K.; Wu, H.M.; Zhang, J.Y.; Zhang, Z.; Zhang, Z.Y.; Rensing, C.; Lin, W.X. Consecutive monoculture regimes differently affected the diversity of the rhizosphere soil viral community and accumulated soil-borne plant viruses. Agric. Ecosyst. Environ. 2022, 337, 108076. [Google Scholar] [CrossRef]
- Waziri, H.M. Plants as antiviral agents. J. Plant Pathol. Microbiol. 2015, 6, 1000254. [Google Scholar] [CrossRef]
- Laufer, M.; Mohammad, H.; Christ, D.S.; Riedel, D.; Maiss, E.; Varrelmann, M.; Liebe, S. Fluorescent labelling of Beet necrotic yellow vein virus and Beet soil-borne mosaic virus for co- and superinfection experiments in Nicotiana benthamiana. J. Gen. Virol. 2018, 99, 1321–1330. [Google Scholar] [CrossRef] [PubMed]
- Abdelkhalek, A.; Salem, M.Z.M.; Hafez, E.; Behiry, S.I.; Qari, S.H. The phytochemical, antifungal, and first report of the antiviral properties of Egyptian Haplophyllum tuberculatum extract. Biology 2020, 9, 248. [Google Scholar] [CrossRef] [PubMed]
- Triolo, E.; Materazzi, A. Rate of tobacco mosaic virus degradation in solarized soil. Riv. Patol. Veg. 1992, 2, 23–32. Available online: https://www.jstor.org/stable/42568064 (accessed on 17 January 2023).
- Yoon, J.B.; Choi, S.K.; Cho, I.S.; Kwon, T.R.; Yang, C.Y.; Seo, M.H.; Yoon, J.Y. Epidemiology of tomato spotted wilt virus in Chrysanthemum morifolium in South Korea and its management using a soil-dwelling predatory mite (Stratiolaelaps scimitus) and essential oils. Virus Res. 2020, 289, 198128. [Google Scholar] [CrossRef]
- Taglienti, A.; Donati, L.; Ferretti, L.; Tomassoli, L.; Sapienza, F.; Sabatino, M.; Di Massimo, G.; Fiorentino, S.; Vecchiarelli, V.; Nota, P.; et al. In vivo antiphytoviral activity of essential oils and hydrosols from Origanum vulgare, Thymus vulgaris, and Rosmarinus officinalis to control zucchini yellow mosaic virus and tomato leaf curl New Delhi virus in Cucurbita pepo L. Front. Microbiol. 2022, 13, 840893. [Google Scholar] [CrossRef]
- Gakuya, D.W.; Itonga, S.M.; Mbaria, J.M.; Muthee, J.K.; Musau, J.K. Ethnobotanical survey of biopesticides and other medicinal plants traditionally used in Meru central district of Kenya. J. Ethnopharmacol. 2013, 145, 547–553. [Google Scholar] [CrossRef]
- Vlassi, A.; Nesler, A.; Perazzolli, M.; Lazazzara, V.; Büschl, C.; Parich, A.; Puopolo, G.; Schuhmacher, R. Volatile organic compounds from Lysobacter capsici AZ78 as potential candidates for biological control of soilborne plant pathogens. Front. Microbiol. 2020, 11, 1748. [Google Scholar] [CrossRef]
- Ajwa, H.A.; Klose, S.; Nelson, S.D.; Minuto, A.; Gullino, M.L. Alternatives to methyl bromide in strawberry production in the United States of America and the Mediterranean region. Phytopathol. Mediterr. 2003, 42, 220–244. [Google Scholar] [CrossRef]
- Mongiano, G.; Zampieri, E.; Morcia, C.; Titone, P.; Volante, A.; Terzi, V.; Tamborini, L.; Valé, G.; Monaco, S. Application of plant-derived bioactive compounds as seed treatments to manage the rice pathogen Fusarium fujikuroi. Crop Prot. 2021, 148, 105739. [Google Scholar] [CrossRef]
- Xie, H.W.; Yan, D.D.; Mao, L.G.; Wang, Q.X.; Li, Y.; Ouyang, C.B.; Guo, M.X.; Cao, A.C. Evaluation of methyl bromide alternatives efficacy against soil-borne pathogens, nematodes and soil microbial community. PLoS ONE 2015, 10, e0117980. [Google Scholar] [CrossRef] [PubMed]
- Verjux, N. Integrated plant protection in potatoes: Review of the French situation. Potato Res. 2017, 60, 307–318. [Google Scholar] [CrossRef]
- Rodrigues, G.C.S.; dos Santos Maia, M.; Cavalcanti, A.B.S.; de Sousa, N.F.; Scotti, M.T.; Scotti, L. In silico studies of Lamiaceae diterpenes with bioinsecticide potential against Aphis gossypii and Drosophila melanogaster. Molecules 2021, 26, 766. [Google Scholar] [CrossRef] [PubMed]
- Arnault, I.; Fleurance, C.; Vey, F.; Fretay, G.D.; Auger, J. Use of Alliaceae residues to control soil-borne pathogens. Ind. Crops Prod. 2013, 49, 265–272. [Google Scholar] [CrossRef]
- Mao, L.G.; Jiang, H.Y.; Zhang, L.; Zhang, Y.N.; Sial, M.U.; Yu, H.T.; Cao, A.C. Replacing methyl bromide with a combination of 1,3-dichloropropene and metam sodium for cucumber production in China. PLoS ONE 2017, 12, e0188137. [Google Scholar] [CrossRef]
- Rajashekar, Y.; Bakthavatsalam, N.; Shivanandappa, T. Botanicals as grain protectants. Psyche 2012, 2012, 646740. [Google Scholar] [CrossRef]
- Ammann, K. Integrated farming: Why organic farmers should use transgenic crops. New Biotechnol. 2008, 25, 101–107. [Google Scholar] [CrossRef]
- Thamsborg, S.M.; Roepstorff, A.; Larsen, M. Integrated and biological control of parasites in organic and conventional production systems. Vet. Parasitol. 1999, 84, 169–186. [Google Scholar] [CrossRef]
- Arunrat, N.; Sereenonchai, S.; Wang, C. Carbon footprint and predicting the impact of climate change on carbon sequestration ecosystem services of organic rice farming and conventional rice farming: A case study in Phichit province, Thailand. J. Environ. Manag. 2021, 289, 112458. [Google Scholar] [CrossRef]
- Hernández-Suárez, E.M.; Beitia, F. Sustainable management methods of orchard insect pests. Insects 2021, 12, 80. [Google Scholar] [CrossRef]
- Katan, J. Physical and cultural methods for the management of soil-borne pathogens. Crop Prot. 2000, 19, 725–731. [Google Scholar] [CrossRef]
- Conti, S.; Villari, G.; Faugno, S.; Melchionna, G.; Somma, S.; Caruso, G. Effects of organic vs. conventional farming system on yield and quality of strawberry grown as an annual or biennial crop in southern Italy. Sci. Hortic. 2014, 180, 63–71. [Google Scholar] [CrossRef]
- Villaverde, J.J.; Sevilla-Morán, B.; Sandín-España, P.; López-Goti, C.; Alonso-Prados, J.L. Challenges of biopesticides under the European Regulation (EC) No. 1107/2009: An overview of new trends in residue analysis. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier: Oxford, UK, 2014; pp. 437–482. [Google Scholar] [CrossRef]
- Smolińska, U.; Kowalska, B. Biological control of the soil-borne fungal pathogen Sclerotinia sclerotiorum—A review. J. Plant Pathol. 2018, 100, 1–12. [Google Scholar] [CrossRef]
- Schmitt, A.; Seddon, B. Biocontrol of plant pathogens with microbial BCAs and plant extracts—Advantages and disadvantages of single and combined use. In Proceedings of the 14th International Reinhardsbrunn Symposium on Modern Fungicides and Antifungal Compounds, Friedrichroda, Germany, 25–29 April 2004. [Google Scholar]
- Barzman, M.; Bàrberi, P.; Birch, A.N.E.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Hommel, B.; Jensen, J.E.; Kiss, J.; Kudsk, P.; et al. Eight principles of integrated pest management. Agron. Sustain. Dev. 2015, 35, 1199–1215. [Google Scholar] [CrossRef]
- Deguine, J.P.; Aubertot, J.N.; Flor, R.J.; Lescourret, F.; Wyckhuys, K.A.G.; Ratnadass, A. Integrated pest management: Good intentions, hard realities. A review. Agron. Sustain. Dev. 2021, 41, 38. [Google Scholar] [CrossRef]
- Kabir, M.H.; Rainis, R. Adoption and intensity of integrated pest management (IPM) vegetable farming in Bangladesh: An approach to sustainable agricultural development. Environ. Dev. Sustain. 2015, 17, 1413–1429. [Google Scholar] [CrossRef]
- Fielding, B.C.; Knowles, C.L.; Vries, F.A.; Klaasen, J.A. Testing of eight medicinal plant extracts in combination with kresoxim-methyl for integrated control of Botrytis cinerea in apples. Agriculture 2015, 5, 400–411. [Google Scholar] [CrossRef] [Green Version]
- Shlevin, E.; Gamliel, A.; Katan, J.; Shtienberg, D. Multi-study analysis of the added benefits of combining soil solarization with fumigants or non-chemical measures. Crop Prot. 2018, 111, 58–65. [Google Scholar] [CrossRef]
- Ait Elallem, B.; Sobeh, M.; Boularbah, A.; Yasri, A. Chemically degraded soil rehabilitation process using medicinal and aromatic plants: Review. Environ. Sci. Pollut. Res. 2021, 28, 73–93. [Google Scholar] [CrossRef]
- Ebadollahi, A.; Ziaee, M.; Palla, F. Essential oils extracted from different species of the Lamiaceae plant family as prospective bioagents against several detrimental pests. Molecules 2020, 25, 1556. [Google Scholar] [CrossRef]
- Posgay, M.; Greff, B.; Kapcsándi, V.; Lakatos, E. Effect of Thymus vulgaris L. essential oil and thymol on the microbiological properties of meat and meat products: A review. Heliyon 2022, 8, e10812. [Google Scholar] [CrossRef] [PubMed]
- Song, B.Z.; Han, Z.H. Assessment of the biocontrol effects of three aromatic plants on multiple trophic levels of the arthropod community in an agroforestry ecosystem. Ecol. Entomol. 2020, 45, 831–839. [Google Scholar] [CrossRef]
- Ehlers, B.K.; Berg, M.P.; Staudt, M.; Holmstrup, M.; Glasius, M.; Ellers, J.; Tomiolo, S.; Madsen, R.B.; Slotsbo, S.; Penuelas, J. Plant secondary compounds in soil and their role in belowground species interactions. Trends Ecol. Evol. 2020, 35, 716–730. [Google Scholar] [CrossRef] [PubMed]
- Thakur, M.; Bhattacharya, S.; Khosla, P.K.; Puri, S. Improving production of plant secondary metabolites through biotic and abiotic elicitation. J. Appl. Res. Med. Aromat. Plants 2019, 12, 1–12. [Google Scholar] [CrossRef]
- Bais, H.P.; Walker, T.S.; Schweizer, H.P.; Vivanco, J.M. Root specific elicitation and antimicrobial activity of rosmarinic acid in hairy root cultures of Ocimum basilicum. Plant Physiol. Biochem. 2002, 40, 983–995. [Google Scholar] [CrossRef]
- Colpas, F.T.; Schwan-Estrada, K.R.F.; Stangarlin, J.R.; Ferrarese, M.D.L.; Scapim, C.A.; Bonaldo, S.M. Induction of plant defense responses by Ocimum gratissimum L. (Lamiaceae) leaf extracts. Summa Phytopathol. 2009, 35, 191–195. [Google Scholar] [CrossRef]
- Zhang, Y.; Han, M.Z.; Song, M.N.; Tian, J.; Song, B.Z.; Hu, Y.J.; Zhang, J.; Yao, Y.C. Intercropping with aromatic plants increased the soil organic matter content and changed the microbial community in a pear orchard. Front. Microbiol. 2021, 12, 616932. [Google Scholar] [CrossRef]
- Borges, D.F.; Lopes, E.A.; Moraes, A.R.F.; Soares, M.S.; Visôtto, L.E.; Oliveira, C.R.; Valente, V.M.M. Formulation of botanicals for the control of plant-pathogens: A review. Crop Prot. 2018, 110, 135–140. [Google Scholar] [CrossRef]
- Mandal, S.M.; Chakraborty, D.; Dey, S. Phenolic acids act as signaling molecules in plant–microbe symbioses. Plant Signal. Behav. 2010, 5, 359–368. [Google Scholar] [CrossRef]
- Misra, P.; Maji, D.; Awasthi, A.; Pandey, S.S.; Yadav, A.; Pandey, A.; Saikia, D.; Babu, C.S.V.; Kalra, A. Vulnerability of soil microbiome to monocropping of medicinal and aromatic plants and its restoration through intercropping and organic amendments. Front. Microbiol. 2019, 10, 2604. [Google Scholar] [CrossRef]
- Mohammadi, H.; Dashi, R.; Farzaneh, M.; Parviz, L.; Hashempour, H. Effects of beneficial root pseudomonas on morphological, physiological, and phytochemical characteristics of Satureja hortensis (Lamiaceae) under water stress. Braz. J. Bot. 2017, 40, 41–48. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Sureda, A.; Tenore, G.C.; Daglia, M.; Sharifi-Rad, M.; Valussi, M.; Tundis, R.; Sharifi-Rad, M.; Loizzo, M.R.; Ademiluyi, A.O.; et al. Biological activities of essential oils: From plant chemoecology to traditional healing systems. Molecules 2017, 22, 70. [Google Scholar] [CrossRef]
- Wenke, K.; Kai, M.; Piechulla, B. Belowground volatiles facilitate interactions between plant roots and soil organisms. Planta 2010, 231, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Parolin, P.; Bresch, C.; Poncet, C.; Desneux, N. Introducing the term ‘Biocontrol Plants’ for integrated pest management. Sci. Agric. 2014, 71, 77–80. [Google Scholar] [CrossRef]
- Saha, A.; Basak, B.B. Scope of value addition and utilization of residual biomass from medicinal and aromatic plants. Ind. Crops Prod. 2020, 145, 111979. [Google Scholar] [CrossRef]
- Zaccardelli, M.; Sorrentino, R.; Caputo, M.; Scotti, R.; De Falco, E.; Pane, C. Stepwise-selected Bacillus amyloliquefaciens and B. subtilis strains from composted aromatic plant waste able to control soil-borne diseases. Agriculture 2020, 10, 30. [Google Scholar] [CrossRef]
- Regnault-Roger, C.; Philogène, B.J.R. Past and current prospects for the use of botanicals and plant allelochemicals in integrated pest management. Pharm. Biol. 2008, 46, 41–52. [Google Scholar] [CrossRef]
- Jacobson, M. Botanical pesticides. Past, present, and future. In Insecticides of Plant Origin; Arnason, J.T., Philogène, B.J.R., Morand, P., Eds.; American Chemical Society: Washington, DC, USA, 1989; pp. 1–10. Available online: https://pubs.acs.org/doi/pdf/10.1021/bk-1989-0387.ch001 (accessed on 17 January 2023).
- Brahmachari, G. Neem—An omnipotent plant: A retrospection. ChemBioChem 2004, 5, 408–421. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Chen, J.B.; Ren, D.; Du, B.; Wu, L.; Zhang, Y.Y.; Wang, Z.Y.; Qian, S. Synthesis of osthol-based botanical fungicides and their antifungal application in crop protection. Bioorg. Med. Chem. 2021, 40, 116184. [Google Scholar] [CrossRef]
- Mulugeta, T.; Muhinyuza, J.B.; Gouws-Meyer, R.; Matsaunyane, L.; Andreasson, E.; Alexandersson, E. Botanicals and plant strengtheners for potato and tomato cultivation in Africa. J. Integr. Agric. 2020, 19, 406–427. [Google Scholar] [CrossRef]
- Asbahani, A.E.; Miladi, K.; Badri, W.; Sala, M.; Addi, E.H.A.; Casabianca, H.; Mousadik, A.E.; Hartmann, D.; Jilale, A.; Renaud, F.N.R.; et al. Essential oils: From extraction to encapsulation. Int. J. Pharm. 2015, 483, 220–243. [Google Scholar] [CrossRef] [PubMed]
- Fierascu, R.C.; Fierascu, I.; Baroi, A.M.; Ortan, A. Selected aspects related to medicinal and aromatic plants as alternative sources of bioactive compounds. Int. J. Mol. Sci. 2021, 22, 1521. [Google Scholar] [CrossRef]
- Ribeiro-Santos, R.; Carvalho-Costa, D.; Cavaleiro, C.; Costa, H.S.; Albuquerque, T.G.; Castilho, M.C.; Ramos, F.; Melo, N.R.; Sanches-Silva, A. A novel insight on an ancient aromatic plant: The rosemary (Rosmarinus officinalis L.). Trends Food Sci. Technol. 2015, 45, 355–368. [Google Scholar] [CrossRef]
- Guesmi, F.; Saidi, I.; Bouzenna, H.; Hfaiedh, N.; Landoulsi, A. Phytocompound variability, antioxidant and antibacterial activities, anatomical features of glandular and aglandular hairs of Thymus hirtus Willd. Ssp. algeriensis Boiss. and Reut. over developmental stages. S. Afr. J. Bot. 2019, 127, 234–243. [Google Scholar] [CrossRef]
- Kundu, A.; Dutta, A.; Mandal, A.; Negi, L.; Malik, M.; Puramchatwad, R.; Antil, J.; Singh, A.; Rao, U.; Saha, S.; et al. A comprehensive in vitro and in silico analysis of nematicidal action of essential oils. Front. Plant Sci. 2021, 11, 614143. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef] [PubMed]
- Mota, I.; Sánchez-Sánchez, J.; Pedro, L.G.; Sousa, M.J. Composition variation of the essential oil from Ocimum basilicum L. cv. Genovese Gigante in response to Glomus intraradices and mild water stress at different stages of growth. Biochem. Syst. Ecol. 2020, 90, 104021. [Google Scholar] [CrossRef]
- Schulz-Bohm, K.; Gerards, S.; Hundscheid, M.; Melenhorst, J.; de Boer, W.; Garbeva, P. Calling from distance: Attraction of soil bacteria by plant root volatiles. ISME J. 2018, 12, 1252–1262. [Google Scholar] [CrossRef]
- Gershenzon, J.; McConkey, M.E.; Croteau, R.B. Regulation of monoterpene accumulation in leaves of peppermint. Plant Physiol. 2000, 122, 205–214. [Google Scholar] [CrossRef]
- van Roon, A.; Parsons, J.R.; te Kloeze, A.M.; Govers, H.A.J. Fate and transport of monoterpenes through soils. Part I. Prediction of temperature dependent soil fate model input-parameters. Chemosphere 2005, 61, 599–609. [Google Scholar] [CrossRef]
- Song, B.Z.; Tang, G.B.; Sang, X.S.; Zhang, J.; Yao, Y.C.; Wiggins, N. Intercropping with aromatic plants hindered the occurrence of Aphis citricola in an apple orchard system by shifting predator–prey abundances. Biocontrol Sci. Technol. 2013, 23, 381–395. [Google Scholar] [CrossRef]
- Vokou, D.; Margaris, N.S.; Lynch, J.M. Effects of volatile oils from aromatic shrubs on soil microorganisms. Soil Biol. Biochem. 1984, 16, 509–513. [Google Scholar] [CrossRef]
- Jouini, A.; Verdeguer, M.; Pinton, S.; Araniti, F.; Palazzolo, E.; Badalucco, L.; Laudicina, V.A. Potential effects of essential oils extracted from Mediterranean aromatic plants on target weeds and soil microorganisms. Plants 2020, 9, 1289. [Google Scholar] [CrossRef] [PubMed]
- Benomari, F.Z.; Andreu, V.; Kotarba, J.; Dib, M.E.A.; Bertrand, C.; Muselli, A.; Costa, J.; Djabou, N. Essential oils from Algerian species of Mentha as new bio-control agents against phytopathogen strains. Environ. Sci. Pollut. Res. 2018, 25, 29889–29900. [Google Scholar] [CrossRef]
- Soković, M.; Tzakou, O.; Pitarokili, D.; Couladis, M. Antifungal activities of selected aromatic plants growing wild in Greece. Nahrung/Food 2002, 46, 317–320. [Google Scholar] [CrossRef]
- Kalemba, D.; Kunicka, A. Antibacterial and antifungal properties of essential oils. Curr. Med. Chem. 2003, 10, 813–829. [Google Scholar] [CrossRef]
- Radaelli, M.; da Silva, B.P.; Weidlich, L.; Hoehne, L.; Flach, A.; da Costa, L.A.M.A.; Ethur, E.M. Antimicrobial activities of six essential oils commonly used as condiments in Brazil against Clostridium perfringens. Braz. J. Microbiol. 2016, 47, 424–430. [Google Scholar] [CrossRef] [Green Version]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential oils’ chemical characterization and investigation of some biological activities: A critical review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef]
- Karpiński, T.M. Essential oils of Lamiaceae family plants as antifungals. Biomolecules 2020, 10, 103. [Google Scholar] [CrossRef]
- Wogiatzi, E.; Gougoulias, N.; Papachatzis, A.; Vagelas, I.; Chouliaras, N. Chemical composition and antimicrobial effects of Greek Origanum species essential oil. Biotechnol. Biotechnol. Equip. 2009, 23, 1322–1324. [Google Scholar] [CrossRef]
- Pitarokili, D.; Tzakou, O.; Kalamarakis, A. Activity of the essential oil of Salvia pomifera L. ssp. calycina (Sm.) Hayek against soil borne pathogens. J. Essent. Oil Res. 2002, 14, 72–75. [Google Scholar] [CrossRef]
- Aguilar-Veloz, L.M.; Calderón-Santoyo, M.; Vázquez González, Y.; Ragazzo-Sánchez, J.A. Application of essential oils and polyphenols as natural antimicrobial agents in postharvest treatments: Advances and challenges. Food Sci. Nutr. 2020, 8, 2555–2568. [Google Scholar] [CrossRef] [PubMed]
- Nieto, G. Biological activities of three essential oils of the Lamiaceae family. Medicines 2017, 4, 63. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, N.; Singh, V.K.; Dwivedy, A.K.; Das, S.; Chaudhari, A.K.; Dubey, N.K. Cistus ladanifer L. essential oil as a plant based preservative against molds infesting oil seeds, aflatoxin B1 secretion, oxidative deterioration and methylglyoxal biosynthesis. LWT 2018, 92, 395–403. [Google Scholar] [CrossRef]
- Sempere-Ferre, F.; Asamar, J.; Castell, V.; Roselló, J.; Santamarina, M.P. Evaluating the antifungal potential of botanical compounds to control Botryotinia fuckeliana and Rhizoctonia solani. Molecules 2021, 26, 2472. [Google Scholar] [CrossRef] [PubMed]
- Marei, G.I.K.; Abdel Rasoul, M.A.; Abdelgaleil, S.A.M. Comparative antifungal activities and biochemical effects of monoterpenes on plant pathogenic fungi. Pestic. Biochem. Physiol. 2012, 103, 56–61. [Google Scholar] [CrossRef]
- Lee, S.O.; Choi, G.J.; Jang, K.S.; Lim, H.K.; Cho, K.Y.; Kim, J.C. Antifungal activity of five plant essential oils as fumigant against postharvest and soilborne plant pathogenic fungi. Plant Pathol. J. 2007, 23, 97–102. [Google Scholar] [CrossRef]
- Bi, Y.; Jiang, H.; Hausbeck, M.K.; Hao, J.J. Inhibitory effects of essential oils for controlling Phytophthora capsici. Plant Dis. 2012, 96, 797–803. [Google Scholar] [CrossRef]
- Laquale, S.; Candido, V.; Avato, P.; Argentieri, M.P.; D’Addabbo, T. Essential oils as soil biofumigants for the control of the root-knot nematode Meloidogyne incognita on tomato. Ann. Appl. Biol. 2015, 167, 217–224. [Google Scholar] [CrossRef]
- Greff, B.; Lakatos, E.; Szigeti, J.; Varga, L. Co-composting with herbal wastes: Potential effects of essential oil residues on microbial pathogens during composting. Crit. Rev. Environ. Sci. Technol. 2021, 51, 457–511. [Google Scholar] [CrossRef]
- Mabrouki, H.; Duarte, C.M.M.; Akretche, D.E. Estimation of total phenolic contents and in vitro antioxidant and antimicrobial activities of various solvent extracts of Melissa officinalis L. Arab. J. Sci. Eng. 2018, 43, 3349–3357. [Google Scholar] [CrossRef]
- Proestos, C.; Sereli, D.; Komaitis, M. Determination of phenolic compounds in aromatic plants by RP-HPLC and GC-MS. Food Chem. 2006, 95, 44–52. [Google Scholar] [CrossRef]
- Costa, D.C.; Costa, H.S.; Albuquerque, T.G.; Ramos, F.; Castilho, M.C.; Sanches-Silva, A. Advances in phenolic compounds analysis of aromatic plants and their potential applications. Trends Food Sci. Technol. 2015, 45, 336–354. [Google Scholar] [CrossRef]
- Seró, R.; Núñez, N.; Núñez, O.; Camprubí, A.; Grases, J.M.; Saurina, J.; Moyano, E.; Calvet, C. Modified distribution in the polyphenolic profile of rosemary leaves induced by plant inoculation with an arbuscular mycorrhizal fungus. J. Sci. Food Agric. 2019, 99, 2966–2973. [Google Scholar] [CrossRef]
- Durazzo, A.; Lucarini, M.; Souto, E.B.; Cicala, C.; Caiazzo, E.; Izzo, A.A.; Novellino, E.; Santini, A. Polyphenols: A concise overview on the chemistry, occurrence, and human health. Phytother. Res. 2019, 33, 2221–2243. [Google Scholar] [CrossRef]
- Sakihama, Y.; Cohen, M.F.; Grace, S.C.; Yamasaki, H. Plant phenolic antioxidant and prooxidant activities: Phenolics-induced oxidative damage mediated by metals in plants. Toxicology 2002, 177, 67–80. [Google Scholar] [CrossRef]
- D’Addabbo, T.; Laquale, S.; Lovelli, S.; Candido, V.; Avato, P. Biocide plants as a sustainable tool for the control of pests and pathogens in vegetable cropping systems. Ital. J. Agron. 2014, 9, 137. [Google Scholar] [CrossRef]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant activity of polyphenols, flavonoids, anthocyanins and carotenoids: Updated review of mechanisms and catalyzing metals. Phytother. Res. 2016, 30, 1379–1391. [Google Scholar] [CrossRef]
- Sytar, O.; Hemmerich, I.; Zivcak, M.; Rauh, C.; Brestic, M. Comparative analysis of bioactive phenolic compounds composition from 26 medicinal plants. Saudi J. Biol. Sci. 2018, 25, 631–641. [Google Scholar] [CrossRef]
- Brown, N.; John, J.A.; Shahidi, F. Polyphenol composition and antioxidant potential of mint leaves. Food Prod. Process. Nutr. 2019, 1, 1. [Google Scholar] [CrossRef]
- Duque-Soto, C.; Borrás-Linares, I.; Quirantes-Piné, R.; Falcó, I.; Sánchez, G.; Segura-Carretero, A.; Lozano-Sánchez, J. Potential antioxidant and antiviral activities of hydroethanolic extracts of selected Lamiaceae species. Foods 2022, 11, 1862. [Google Scholar] [CrossRef] [PubMed]
- Mojzer, E.B.; Hrnčič, M.K.; Škerget, M.; Knez, Ž.; Bren, U. Polyphenols: Extraction methods, antioxidative action, bioavailability and anticarcinogenic effects. Molecules 2016, 21, 901. [Google Scholar] [CrossRef] [PubMed]
- Bouarab-Chibane, L.; Forquet, V.; Lantéri, P.; Clément, Y.; Léonard-Akkari, L.; Oulahal, N.; Degraeve, P.; Bordes, C. Antibacterial properties of polyphenols: Characterization and QSAR (Quantitative Structure–Activity Relationship) models. Front. Microbiol. 2019, 10, 829. [Google Scholar] [CrossRef] [PubMed]
- Proestos, C.; Chorianopoulos, N.; Nychas, G.J.E.; Komaitis, M. RP-HPLC analysis of the phenolic compounds of plant extracts. Investigation of their antioxidant capacity and antimicrobial activity. J. Agric. Food Chem. 2005, 53, 1190–1195. [Google Scholar] [CrossRef]
- Cetin-Karaca, H.; Newman, M.C. Antimicrobial efficacy of plant phenolic compounds against Salmonella and Escherichia coli. Food Biosci. 2015, 11, 8–16. [Google Scholar] [CrossRef]
- Daglia, M. Polyphenols as antimicrobial agents. Curr. Opin. Biotechnol. 2012, 23, 174–181. [Google Scholar] [CrossRef]
- Shalaby, S.; Horwitz, B.A. Plant phenolic compounds and oxidative stress: Integrated signals in fungal–plant interactions. Curr. Genet. 2015, 61, 347–357. [Google Scholar] [CrossRef]
- Gao, X.Z.; Xu, Z.J.; Liu, G.T.; Wu, J. Polyphenols as a versatile component in tissue engineering. Acta Biomater. 2021, 119, 57–74. [Google Scholar] [CrossRef]
- Khameneh, B.; Iranshahy, M.; Soheili, V.; Bazzaz, B.S.F. Review on plant antimicrobials: A mechanistic viewpoint. Antimicrob. Resist. Infect. Control 2019, 8, 118. [Google Scholar] [CrossRef]
- Summers, C.B.; Felton, G.W. Prooxidant effects of phenolic acids on the generalist herbivore Helicoverpa zea (Lepidoptera: Noctuidae): Potential mode of action for phenolic compounds in plant anti-herbivore chemistry. Insect Biochem. Mol. Biol. 1994, 24, 943–953. [Google Scholar] [CrossRef]
- Simmonds, M.S.J. Flavonoid–insect interactions: Recent advances in our knowledge. Phytochemistry 2003, 64, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Widmer, T.L.; Laurent, N. Plant extracts containing caffeic acid and rosmarinic acid inhibit zoospore germination of Phytophthora spp. pathogenic to Theobroma cacao. Eur. J. Plant Pathol. 2006, 115, 377–388. [Google Scholar] [CrossRef]
- Al-Huqail, A.A.; Behiry, S.I.; Salem, M.Z.M.; Ali, H.M.; Siddiqui, M.H.; Salem, A.Z.M. Antifungal, antibacterial, and antioxidant activities of Acacia saligna (Labill.) H. L. Wendl. flower extract: HPLC analysis of phenolic and flavonoid compounds. Molecules 2019, 24, 700. [Google Scholar] [CrossRef] [PubMed]
- Matsubara, Y.; Yamashita, Y.; Liu, J. Antifungal and antioxidative ability in Lamiaceae herbs. Acta Hortic. 2015, 1105, 109–114. [Google Scholar] [CrossRef]
- Ahmad, H.; Matsubara, Y. Suppression of Fusarium wilt in Cyclamen by using sage water extract and identification of antifungal metabolites. Australas. Plant Pathol. 2020, 49, 213–220. [Google Scholar] [CrossRef]
- López, V.; Akerreta, S.; Casanova, E.; García-Mina, J.M.; Cavero, R.Y.; Calvo, M.I. In vitro antioxidant and anti-rhizopus activities of Lamiaceae herbal extracts. Plant Foods Hum. Nutr. 2007, 62, 151–155. [Google Scholar] [CrossRef]
- Skrzypek, E.; Repka, P.; Stachurska-Swakoń, A.; Barabasz-Krasny, B.; Możdżeń, K. Allelopathic effect of aqueous extracts from the leaves of peppermint (Mentha piperita L.) on selected physiological processes of common sunflower (Helianthus annuus L.). Not. Bot. Horti Agrobot. 2015, 43, 335–342. [Google Scholar] [CrossRef] [Green Version]
- Isman, M.B. Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–66. [Google Scholar] [CrossRef]
- NÉBIH Fertilizer Database. Available online: https://termesnovelo.nebih.gov.hu/Engedelykereso/kereso (accessed on 17 January 2023).
- NÉBIH Pesticide Database. Available online: https://novenyvedoszer.nebih.gov.hu/Engedelykereso/kereso (accessed on 17 January 2023).
- EU Pesticide Database. Available online: https://food.ec.europa.eu/plants/pesticides/eu-pesticides-database_en (accessed on 17 January 2023).
- Lin, C.; Owen, S.M.; Peñuelas, J. Volatile organic compounds in the roots and rhizosphere of Pinus spp. Soil Biol. Biochem. 2007, 39, 951–960. [Google Scholar] [CrossRef]
- Li, X.G.; de Boer, W.; Zhang, Y.A.; Ding, C.F.; Zhang, T.L.; Wang, X.X. Suppression of soil-borne Fusarium pathogens of peanut by intercropping with the medicinal herb Atractylodes lancea. Soil Biol. Biochem. 2018, 116, 120–130. [Google Scholar] [CrossRef]
- Mamolos, A.P.; Kalburtji, K.L. Significance of allelopathy in crop rotation. J. Crop Prod. 2001, 4, 197–218. [Google Scholar] [CrossRef]
- Verma, R.K.; Yadav, A.; Rahman, L.U.; Kalra, A.; Patra, D.D. Influence the status of soil chemical and biological properties by intercropping. Int. J. Recycl. Org. Waste Agric. 2014, 3, 46. [Google Scholar] [CrossRef]
- Vukicevich, E.; Lowery, T.; Bowen, P.; Úrbez-Torres, J.R.; Hart, M. Cover crops to increase soil microbial diversity and mitigate decline in perennial agriculture. A review. Agron. Sustain. Dev. 2016, 36, 48. [Google Scholar] [CrossRef]
- Khan, K.; Tewari, S.K.; Semwal, M.; Verma, R.K. Effects of aromatic crops based cropping systems and residue returning on soil CO2 emission compared to fallow soil. J. Appl. Res. Med. Aromat. Plants 2022, 26, 100351. [Google Scholar] [CrossRef]
- Khan, K.; Verma, R.K. Diversifying cropping systems with aromatic crops for better productivity and profitability in subtropical north Indian plains. Ind. Crops Prod. 2018, 115, 104–110. [Google Scholar] [CrossRef]
- Raveau, R.; Fontaine, J.; Hijri, M.; Sahraoui, A.L.H. The aromatic plant clary sage shaped bacterial communities in the roots and in the trace element-contaminated soil more than mycorrhizal inoculation—A two-year monitoring field trial. Front. Microbiol. 2020, 11, 586050. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.L.; Liu, X.Y.; Zhou, C.; Han, S.F.; Xu, F.R.; Dong, X. Root-associated microbiomes of Panax notoginseng under the combined effect of plant development and Alpinia officinarum Hance essential oil. Molecules 2022, 27, 6014. [Google Scholar] [CrossRef] [PubMed]
- Bafana, A. Diversity and metabolic potential of culturable root-associated bacteria from Origanum vulgare in sub-Himalayan region. World J. Microbiol. Biotechnol. 2013, 29, 63–74. [Google Scholar] [CrossRef]
- Karthikeyan, B.; Jaleel, C.A.; Lakshmanan, G.M.A.; Deiveekasundaram, M. Studies on rhizosphere microbial diversity of some commercially important medicinal plants. Colloids Surf. B Biointerfaces 2008, 62, 143–145. [Google Scholar] [CrossRef]
- Adamović, D.; Đalović, I.; Mrkovački, N. Microbial abundance in rhizosphere of medicinal and aromatic plant species in conventional and organic growing systems. Ratar. Povrt. 2015, 52, 1–6. [Google Scholar] [CrossRef]
- Jisha, C.K.; Bauddh, K.; Shukla, S.K. Phytoremediation and bioenergy production efficiency of medicinal and aromatic plants. In Phytoremediation Potential of Bioenergy Plants; Bauddh, K., Singh, B., Korstad, J., Eds.; Springer: Singapore, 2017; pp. 287–304. [Google Scholar] [CrossRef]
- Chen, L.J.; Wu, X.Q.; Xu, Y.; Wang, B.L.; Liu, S.; Niu, J.F.; Wang, Z. Microbial diversity and community structure changes in the rhizosphere soils of Atractylodes lancea from different planting years. Plant Signal. Behav. 2021, 16, 1854507. [Google Scholar] [CrossRef] [PubMed]
- Karkanis, A.; Alexiou, A.; Katsaros, C.; Petropoulos, S. Allelopathic activity of spearmint (Mentha spicata L.) and peppermint (Mentha × piperita L.) reduces yield, growth, and photosynthetic rate in a succeeding crop of maize (Zea mays L.). Agronomy 2019, 9, 461. [Google Scholar] [CrossRef]
- Dittrich, F.; Iserloh, T.; Treseler, C.H.; Hüppi, R.; Ogan, S.; Seeger, M.; Thiele-Bruhn, S. Crop diversification in viticulture with aromatic plants: Effects of intercropping on grapevine productivity in a steep-slope vineyard in the Mosel area, Germany. Agriculture 2021, 11, 95. [Google Scholar] [CrossRef]
- Chand, S.; Singh, D.; Madegowda, M.; Sikka, A. Medicinal and aromatic plants for soil and water conservation in Nilgiris, Tamil Nadu, India: An economic analysis. Mol. Soil Biol. 2015, 665, 1–6. [Google Scholar] [CrossRef]
- Dhima, K.V.; Vasilakoglou, I.B.; Gatsis, T.D.; Panou-Philotheou, E.; Eleftherohorinos, I.G. Effects of aromatic plants incorporated as green manure on weed and maize development. Field Crops Res. 2009, 110, 235–241. [Google Scholar] [CrossRef]
- Kamil, D.; Kumar, R.; Sinha, A. Effect of green manuring of Crotolaria juncea L. on some soil-borne pathogens. Indian Phytopathol. 2009, 62, 304–309. [Google Scholar]
- Dojima, T.; Craker, L.E. Potential benefits of soil microorganisms on medicinal and aromatic plants. In Medicinal and Aromatic Crops: Production, Phytochemistry, and Utilization; Jeliazkov, V.D., Cantrell, C.L., Eds.; American Chemical Society: Washington, DC, USA, 2016; pp. 75–90. [Google Scholar] [CrossRef]
- Kushwaha, R.K.; Rodrigues, V.; Kumar, V.; Patel, H.; Raina, M.; Kumar, D. Soil microbes–medicinal plants interactions: Ecological diversity and future prospect. In Plant Microbe Symbiosis; Varma, A., Tripathi, S., Prasad, R., Eds.; Springer: Cham, Switzerland, 2020; pp. 263–286. [Google Scholar] [CrossRef]
- Castronovo, L.M.; Calonico, C.; Ascrizzi, R.; Del Duca, S.; Delfino, V.; Chioccioli, S.; Vassallo, A.; Strozza, I.; De Leo, M.; Biffi, S.; et al. The cultivable bacterial microbiota associated to the medicinal plant Origanum vulgare L.: From antibiotic resistance to growth-inhibitory properties. Front. Microbiol. 2020, 11, 862. [Google Scholar] [CrossRef]
- Tiwari, S.; Pandey, R.; Gross, A. Identification of rhizospheric microorganisms that manages root knot nematode and improve oil yield in sweet basil (Ocimum basilicum L.). Agronomy 2021, 11, 570. [Google Scholar] [CrossRef]
- Chowdhary, K.; Kaushik, N. Biodiversity study and potential of fungal endophytes of peppermint and effect of their extract on chickpea rot pathogens. Arch. Phytopathol. Plant Prot. 2018, 51, 139–155. [Google Scholar] [CrossRef]
- Rao, E.V.S.P. Medicinal and aromatic plants for crop diversification and their agronomic implications. Indian J. Agron. 2009, 54, 215–220. [Google Scholar]
- Ahmad, H.; Kobayashi, M.; Matsubara, Y. Changes in secondary metabolites and free amino acid content in tomato with Lamiaceae herbs companion planting. Am. J. Plant Sci. 2020, 11, 1878–1889. [Google Scholar] [CrossRef]
- Parolin, P.; Bresch, C.; Poncet, C.; Suay-Cortez, R.; Van Oudenhove, L. Testing basil as banker plant in IPM greenhouse tomato crops. Int. J. Pest Manag. 2015, 61, 235–242. [Google Scholar] [CrossRef]
- Carvalho, L.M.; Nunes, M.U.C.; Oliveira, I.R.; Leal, M.L.S. Produtividade do tomateiro em cultivo solteiro e consorciado com espécies aromáticas e medicinais. Hortic. Bras. 2009, 27, 458–464. [Google Scholar] [CrossRef]
- Puig, C.G.; Álvarez-Iglesias, L.; Reigosa, M.J.; Pedrol, N. Eucalyptus globulus leaves incorporated as green manure for weed control in maize. Weed Sci. 2013, 61, 154–161. [Google Scholar] [CrossRef]
- Thakur, M.; Kumar, R. Microclimatic buffering on medicinal and aromatic plants: A review. Ind. Crops Prod. 2021, 160, 113144. [Google Scholar] [CrossRef]
- de Elguea-Culebras, G.O.; Bravo, E.M.; Sánchez-Vioque, R. Potential sources and methodologies for the recovery of phenolic compounds from distillation residues of Mediterranean aromatic plants. An approach to the valuation of by-products of the essential oil market—A review. Ind. Crops Prod. 2022, 175, 114261. [Google Scholar] [CrossRef]
- Skendi, A.; Irakli, M.; Chatzopoulou, P.; Bouloumpasi, E.; Biliaderis, C.G. Phenolic extracts from solid wastes of the aromatic plant essential oil industry: Potential uses in food applications. Food Chem. Adv. 2022, 1, 100065. [Google Scholar] [CrossRef]
- Basak, B.B. Recycling of waste biomass and mineral powder for preparation of potassium-enriched compost. J. Mater. Cycles Waste Manag. 2018, 20, 1409–1415. [Google Scholar] [CrossRef]
- Yohalem, D.; Passey, T. Amendment of soils with fresh and post-extraction lavender (Lavandula angustifolia) and lavandin (Lavandula × intermedia) reduce inoculum of Verticillium dahliae and inhibit wilt in strawberry. Appl. Soil Ecol. 2011, 49, 187–196. [Google Scholar] [CrossRef]
- Kadoglidou, K.; Chalkos, D.; Karamanoli, K.; Eleftherohorinos, I.G.; Constantinidou, H.I.A.; Vokou, D. Aromatic plants as soil amendments: Effects of spearmint and sage on soil properties, growth and physiology of tomato seedlings. Sci. Hortic. 2014, 179, 25–35. [Google Scholar] [CrossRef]
- Klein, E.; Katan, J.; Gamliel, A. Combining residues of herb crops with soil heating for control of soilborne pathogens in a controlled laboratory system. Crop Prot. 2011, 30, 368–374. [Google Scholar] [CrossRef]
- Vokou, D.; Chalkos, D.; Karamanlidou, G.; Yiangou, M. Activation of soil respiration and shift of the microbial population balance in soil as a response to Lavandula stoechas essential oil. J. Chem. Ecol. 2002, 28, 755–768. [Google Scholar] [CrossRef] [PubMed]
- Gravanis, F.; Chouliaras, N.; Vagelas, I.; Gougoulias, N. The effect of oregano (Origanum vulgare) as an alternative soil-borne pathogen control agent, on soil organic matter biodegradation and other soil chemical properties. In Proceedings of the BCPC International Congress on Crop Science & Technology, Glasgow, UK, 31 October–2 November 2005. [Google Scholar]
- Chouliaras, N.; Gravanis, F.; Vasilakoglou, I.; Gougoulias, N.; Vagelas, I.; Kapotis, T.; Wogiatzi, E. The effect of basil (Ocimum basilicum L.) on soil organic matter biodegradation and other soil chemical properties. J. Sci. Food Agric. 2007, 87, 2416–2419. [Google Scholar] [CrossRef]
- Ainalidou, A.; Bouzoukla, F.; Menkissoglu-Spiroudi, U.; Vokou, D.; Karamanoli, K. Impacts of decaying aromatic plants on the soil microbial community and on tomato seedling growth and metabolism: Suppression or stimulation? Plants 2021, 10, 1848. [Google Scholar] [CrossRef]
- Argyropoulou, M.D.; Karmezi, M.; Tsiafouli, M.; Chalkos, D.; Bountla, A.; Vokou, D. Soil amendments with spearmint, peppermint and rosemary enhance the community of free-living nematodes and improve soil quality, while having strikingly different effects on plant growth. Life 2022, 12, 1121. [Google Scholar] [CrossRef]
- Deka, H.; Deka, S.; Baruah, C.K.; Das, J.; Hoque, S.; Sarma, H.; Sarma, N.S. Vermicomposting potentiality of Perionyx excavatus for recycling of waste biomass of java citronella—An aromatic oil yielding plant. Bioresour. Technol. 2011, 102, 11212–11217. [Google Scholar] [CrossRef]
- El-Helw, N.O.; El-Gendy, A.O.; El-Gebaly, E.; Hassan, H.M.; Rateb, M.E.; El-Nesr, K.A. Characterization of natural bioactive compounds produced by isolated bacteria from compost of aromatic plants. J. Appl. Microbiol. 2019, 126, 443–451. [Google Scholar] [CrossRef]
- Pane, C.; Piccolo, A.; Spaccini, R.; Celano, G.; Villecco, D.; Zaccardelli, M. Agricultural waste-based composts exhibiting suppressivity to diseases caused by the phytopathogenic soil-borne fungi Rhizoctonia solani and Sclerotinia minor. Appl. Soil Ecol. 2013, 65, 43–51. [Google Scholar] [CrossRef]
- Pandey, R.; Kalra, A. Inhibitory effects of vermicompost produced from agro-waste of medicinal and aromatic plants on egg hatching in Meloidogyne incognita (Kofoid and White) Chitwood. Curr. Sci. 2010, 98, 833–835. [Google Scholar]
- Singh, R.; Soni, S.K.; Awasthi, A.; Kalra, A. Evaluation of vermicompost doses for management of root-rot disease complex in Coleus forskohlii under organic field conditions. Australas. Plant Pathol. 2012, 41, 397–403. [Google Scholar] [CrossRef]
- Zhou, Y.; Selvam, A.; Wong, J.W.C. Effect of Chinese medicinal herbal residues on microbial community succession and anti-pathogenic properties during co-composting with food waste. Bioresour. Technol. 2016, 217, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Bajer, T.; Šilha, D.; Ventura, K.; Bajerová, P. Composition and antimicrobial activity of the essential oil, distilled aromatic water and herbal infusion from Epilobium parviflorum Schreb. Ind. Crops Prod. 2017, 100, 95–105. [Google Scholar] [CrossRef]
- Ozturk, I.; Tornuk, F.; Caliskan-Aydogan, O.; Durak, M.Z.; Sagdic, O. Decontamination of iceberg lettuce by some plant hydrosols. LWT 2016, 74, 48–54. [Google Scholar] [CrossRef]
- Tabti, L.; Dib, M.E.A.; Djabou, N.; Benyelles, N.G.; Paolini, J.; Costa, J.; Muselli, A. Control of fungal pathogens of Citrus sinensis L. by essential oil and hydrosol of Thymus capitatus L. J. Appl. Bot. Food Qual. 2014, 87, 279–285. [Google Scholar] [CrossRef]
- Gaspar-Pintiliescu, A.; Mihai, E.; Stanciuc, A.M.; Anton, R.E.; Popescu, A.F.; Craciunescu, O. Polyphenolic content, antioxidant and antimicrobial activity of hydrosols from some aromatic Lamiaceae plants. Proceedings 2020, 57, 21. [Google Scholar] [CrossRef]
- Andrés, M.F.; González-Coloma, A.; Muñoz, R.; De la Peña, F.; Julio, L.F.; Burillo, J. Nematicidal potential of hydrolates from the semi industrial vapor-pressure extraction of Spanish aromatic plants. Environ. Sci. Pollut. Res. 2018, 25, 29834–29840. [Google Scholar] [CrossRef] [PubMed]
- Ntalli, N.; Parlapani, A.B.; Tzani, K.; Samara, M.; Boutsis, G.; Dimou, M.; Menkissoglu-Spiroudi, U.; Monokrousos, N. Thymus citriodorus (Schreb) botanical products as ecofriendly nematicides with bio-fertilizing properties. Plants 2020, 9, 202. [Google Scholar] [CrossRef] [Green Version]
- Sainz, P.; Andrés, M.F.; Martínez-Díaz, R.A.; Bailén, M.; Navarro-Rocha, J.; Díaz, C.E.; González-Coloma, A. Chemical composition and biological activities of Artemisia pedemontana subsp. assoana essential oils and hydrolate. Biomolecules 2019, 9, 558. [Google Scholar] [CrossRef]
- Pino-Otín, R.; Val, J.; Ballestero, D.; Navarro, E.; Sánchez, E.; Mainar, A.M. Impact of Artemisia absinthium hydrolate extracts with nematicidal activity on non-target soil organisms of different trophic levels. Ecotoxicol. Environ. Saf. 2019, 180, 565–574. [Google Scholar] [CrossRef]
Figure 1.
Aromatic and medicinal plants as part of non-chemical plant protections systems: effects and methods of application. The icons in circles were made by Blackwoods, Freepik, Kerismaker, and Surang (www.flaticon.com). The plant illustration on the left is based on the work of Mariya Kazakova (purchased from www.shutterstock.com).
Figure 1.
Aromatic and medicinal plants as part of non-chemical plant protections systems: effects and methods of application. The icons in circles were made by Blackwoods, Freepik, Kerismaker, and Surang (www.flaticon.com). The plant illustration on the left is based on the work of Mariya Kazakova (purchased from www.shutterstock.com).

Figure 2.
(a) Post-extraction lavender (Lavandula angustifolia) waste; (b) Antifungal effect of lavender waste extracts on phytopathogenic Sclerotinia sclerotiorum. Photographs taken by Babett Greff.
Figure 2.
(a) Post-extraction lavender (Lavandula angustifolia) waste; (b) Antifungal effect of lavender waste extracts on phytopathogenic Sclerotinia sclerotiorum. Photographs taken by Babett Greff.


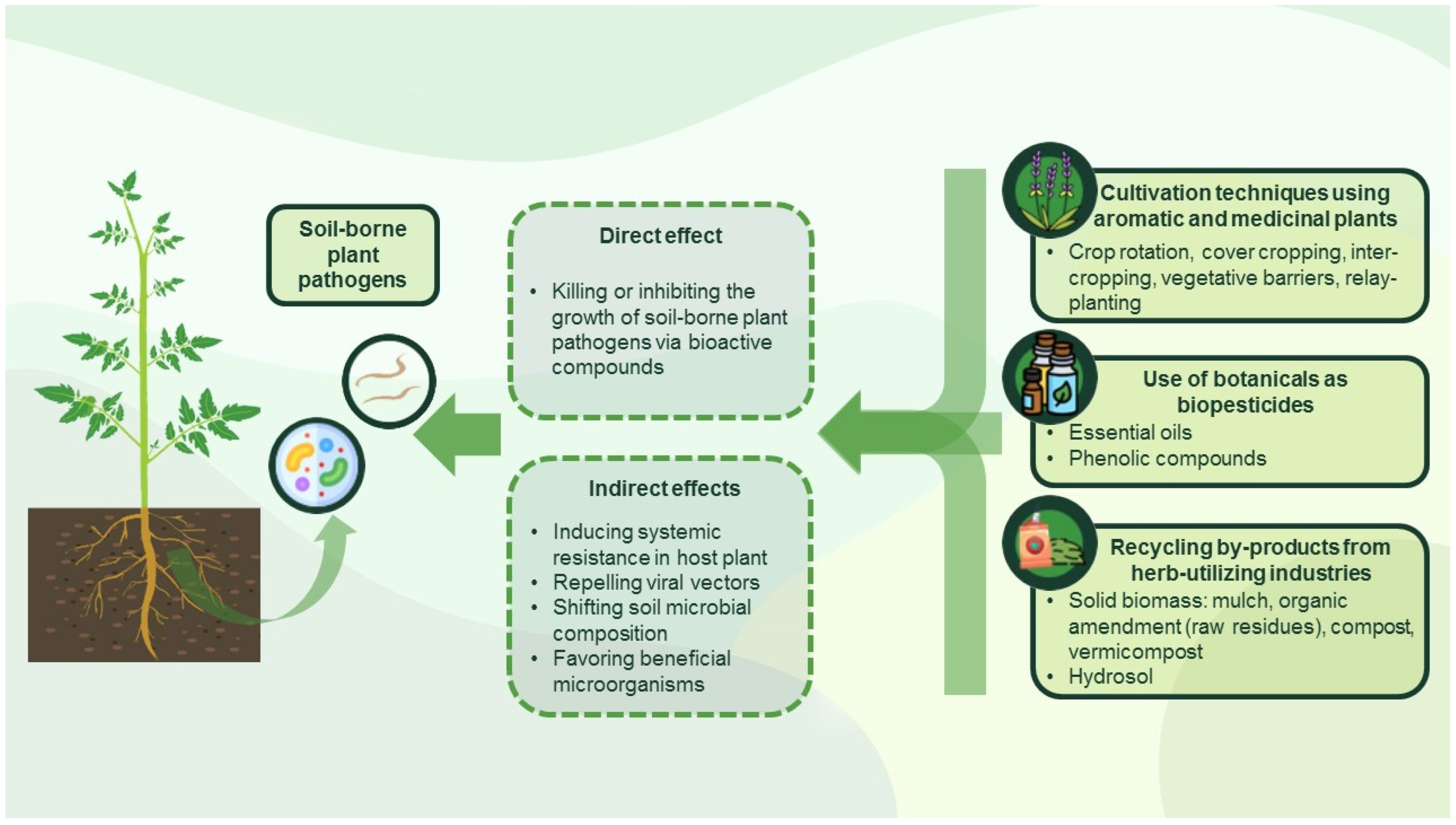{kind=link}
{kind=link}
Table 1.
Properties of essential oils and/or essential oil constituents, dealt with in this review paper, against soil-borne organisms and related references.
Table 1.
Properties of essential oils and/or essential oil constituents, dealt with in this review paper, against soil-borne organisms and related references.
| Essential Oil (EO) | Target Organism | Effect | Reference |
|---|---|---|---|
| Thymus pallescens, Artemisia herba-alba Asso, Laurus nobilis L., Cymbopogon citratus (de Candolle ex Nees) Stapf. | Fusarium oxysporum f. sp. ciceris Padwick | Inhibition of mycelial growth, sporulation, and spore germination | [46] |
| Mentha piperita L., Thymus vulgaris L., Lavandula angustifolia Mill. | Verticillium dahliae Kleb. isolates | Inhibition of mycelial growth | [51] |
| Origanum dictamnus, Mentha pulegium, Origanum vulgare, Melissa officinalis | Meloidogyne incognita | High nematicidal activity (irreversible motility loss of nematode juveniles) | [65] |
| Satureja thymbra, Rosmarinus officinalis | Penicillium citrinum | Suppression of spore germination | [139] |
| Satureja thymbra | Mucor hiemalis | Complete inhibition of mycelial growth | [139] |
| Mentha spp. | Penicillium expansum | Inhibition of mycelial growth | [141] |
| Origanum onites L., Salvia thymbra L. | Alternaria alternata, Aspergillus spp., Penicillium spp., Trichoderma viride, Cladosporium cladosporioides, Phomopsis helianthi | Strong antifungal activity against the microorganisms tested | [142] |
| Origanum spp. | Verticillium dahliae, Fusarium oxysporum f. sp. lycopersici, Sclerotinia sclerotiorum, Pythium spp. | Inhibition of mycelial growth | [147] |
| Salvia pomifera L. ssp. calycina (Sm.) Hayek | Fusarium oxysporum f. sp. dianthi, Fusarium solani f. sp. cucurbitae, Fusarium proliferatum, Verticillium dahliae, Sclerotinia sclerotiorum, Rhizoctonia solani | Inhibition of mycelial growth | [148] |
| EO constituents (carvacrol, cinnamaldehyde, eugenol, thymol) | Rhizoctonia solani | Inhibition of mycelial growth | [152] |
| EO constituents [camphene, (R)-camphor, (R)-carvone, 1,8-cineole, cuminaldehyde, (S)-fenchone, geraniol, (R)-linalool, (1R,2S,5R)-menthol, myrcene] | Aspergillus niger, Fusarium oxysporum, Penicillium digitatum, Rhizoctonia solani | Inhibition of mycelial growth | [153] |
| EO constituents [thymol, (S)-limonene] | Inhibition of mycelial growth, inhibitory effect on pectin methyl esterase and cellulase | ||
| Cuminum cyminum, Eucalyptus citriodora, Origanum vulgare | Fusarium oxysporum, Pythium ultimum, Rhizoctonia solani | Inhibition of mycelial growth | [154] |
| Origanum syraicum, Cymbopogon martini, Thymus vulgaris | Phytophthora capsici isolates | Strong antifungal activity, control of production and germination of sporangia and zoospores, oospore production, and mycelial growth reduction of Phytophthora capsici population in soil, suppression of infection on zucchini fruits | [155] |
Table 2.
Properties of extracts from aromatic and medicinal plants, dealt with in this review paper, against soil-borne organisms and related references.
Table 2.
Properties of extracts from aromatic and medicinal plants, dealt with in this review paper, against soil-borne organisms and related references.
| Plant Extract | Type of the Extract | Target Organism | Effect | Reference |
|---|---|---|---|---|
| Mentha piperita L., Thymus vulgaris L., Lavandula angustifolia Mill. | - | Verticillium dahliae Kleb. isolates | Inhibition of mycelial growth | [51] |
| Orginum vulgare | Hydrosol from EO distillation | Meloidogyne javanica | Nematostatic/nematicidal activity (paralysis of second-stage juveniles) | [64] |
| Orginum vulgare, Thymus citriodorus | Water extract of plant powders | Meloidogyne incognita, Meloidogyne javanica | ||
| Rosmarinus officinalis, Lavandula angustifolia, Lavandula angustifolia × Lavandula spicata, Salvia officinalis | Hot water extract of leaves | Phytophthora spp. isolates | Control of zoospore germination | [179] |
| Acacia saligna | Water extract of flowers | Fusarium culmorum, Rhizoctonia solani, Penicillium chrysogenum | Inhibition of mycelial growth | [180] |
| Acacia saligna | Water extract of flowers | Agrobacterium tumefaciens, Pectobacterium carotovorum subsp. carotovorum | Inhibition of bacterial growth | [180] |
| Origanum vulgare L., Salvia officinalis L., Melissa officinalis L. | Water extract of shoots | Fusarium oxysporum f. sp. asparagi | Suppression of fungal propagation | [181] |
| Salvia officinalis L. | Water extract of shoots | Fusarium oxysporum f. sp. cyclaminis | Suppression of fungal pathogen, reduction of disease indices in cyclamen roots and shoots | [182] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Greff, B.; Sáhó, A.; Lakatos, E.; Varga, L. Biocontrol Activity of Aromatic and Medicinal Plants and Their Bioactive Components against Soil-Borne Pathogens. Plants 2023, 12, 706. https://doi.org/10.3390/plants12040706
AMA Style
Greff B, Sáhó A, Lakatos E, Varga L. Biocontrol Activity of Aromatic and Medicinal Plants and Their Bioactive Components against Soil-Borne Pathogens. Plants. 2023; 12(4):706. https://doi.org/10.3390/plants12040706
Chicago/Turabian StyleGreff, Babett, András Sáhó, Erika Lakatos, and László Varga. 2023. "Biocontrol Activity of Aromatic and Medicinal Plants and Their Bioactive Components against Soil-Borne Pathogens" Plants 12, no. 4: 706. https://doi.org/10.3390/plants12040706
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.